


近年来,农作物遭受生物和非生物胁迫压力越来越大,严重制约了农业发展。研究[1-
本研究基于花生全基因组数据,系统鉴定了AhFKBP基因家族成员,并对其系统进化、基因结构、染色体分布、共线性关系、顺式作用元件及表达模式等进行分析,旨在揭示FKBP基因在花生生长发育和逆境响应中的潜在功能,为进一步研究花生抗逆分子机制及遗传改良提供理论依据。
1 材料与方法
1.1 花生FKBP基因的鉴定与特征分析
以Pfam数据库中的PF00254保守结构域文件对花生基因组进行检索;同时,以拟南芥和水稻的FKBP蛋白序列为种子序列,通过BLASTP方法在花生基因组数据库(Peanut Base)中搜索花生FKBP蛋白序列,以交集方式合并两部分的鉴定结果。随后利用鉴定得到的蛋白质序列初步构建花生特异性的FKBP HMM文件,并通过该文件对花生基因组数据库再次检索(E-value<1e-2),最终经SMART(
运用ExPASy(
1.2 花生FKBP基因家族系统进化树构建与分析
在TAIR(
1.3 花生FKBP基因家族保守结构分析
根据花生基因组的注释信息,使用biolinux系统将花生gf3文件转换为gtf文件,获取FKBP基因的UTR区、外显子和内含子结构信息。利用MEME网站(
1.4 花生FKBP基因家族染色体定位及共线性分析
利用TBtools中的Gtf/Gf3序列提取工具获取花生基因的所有CDS(编码序列),并通过批量CDS转蛋白质工具(Batch Translate CDS to Protein)将CDS序列翻译为蛋白质序列。通过BLAST比对筛选FKBP基因家族的串联重复和片段重复事件。结合花生基因组的染色体位置信息,使用TBtools的基因定位可视化工具和高级Circos工具,绘制该基因家族的染色体定位图及Circos环形图。
利用花生基因组注释信息获得AhFKBP基因的染色体位置,使用TBtools软件绘制AhFKBP基因在染色体上的分布图。利用BIO-LINUX系统,通过双向BLAST比较和MCScanx工具进一步分析AhFKBP基因在花生不同染色体间的共线性。
1.5 花生与4种代表植物FKBP基因的种间共线性分析
从EnsemblPlants数据库中下载花生、黄瓜、小麦、水稻和拟南芥的蛋白质序列和gf3文件。使用TBtools软件中的BLAST工具进行双向比对,运行MCscanX Wrapper以获取所有基因的位置信息和共线性区块(最小区块大小设为30)。利用TBtools软件中的多重共线性绘图工具绘制花生与黄瓜、小麦、水稻和拟南芥的共线性比较图谱。
1.6 花生FKBP基因家族顺式作用元件分析
利用TBtools提取AhFKBP基因上游2000 bp启动子序列,将启动子序列上传到Plant CARE(
1.7 花生FKBP基因组的表达分析
在Peanut Base中下载花生不同组织的转录组表达数据,在转录组数据中检索花生FKBP基因家族,利用TBtools软件进行热图绘制,分析花生FKBP基因的表达模式。
2 结果与分析
2.1 花生FKBP基因家族的鉴定分析
以拟南芥和水稻的FKBP蛋白为种子序列在花生基因组数据库(Peanut Base)搜索花生FKBP蛋白序列并构建花生特异性FKBP HMM文件,共得到56个AhFKBP基因家族成员(表1)。花生FKBP基因家族的CDS长度为339~2841 bp,氨基酸数为112~946个,分子量为12.00~105.08 kDa,其中AhFKBP24最小,AhFKBP7最大;等电点范围为4.83(AhFKBP16)~9.24(AhFKBP8),其中32个基因蛋白质等电点<7.00,24个基因蛋白质等电点>7.00;鉴定出AhFKBP10、AhFKBP18、AhFKBP19、AhFKBP42和AhFKBP47均具有1个可预测的跨膜结构域;不稳定系数范围为19.82(AhFKBP39)~60.07(AhFKBP4),脂肪系数为57.89(AhFKBP15)~111.62(AhFKBP32);AhFKBP3、AhFKBP5和AhFKBP32平均疏水性>0,其余基因均<0,表明花生FKBP蛋白具有较强的亲水性。对AhFKBP蛋白进行亚细胞定位预测,发现11个蛋白定位于叶绿体上,1个蛋白定位于叶绿体外膜,21个蛋白定位于细胞质,7个定位于细胞外间隙中,3个定位于线粒体中,12个定位于细胞核中,1个定位于细胞器膜上。
表1 AhFKBP基因家族成员及其理化性质
Table 1
| 基因名称 Gene name | 基因位点 Gene locus | CDS长度 CDS length (bp) | 氨基酸数 Number of amino acids | 分子量 Molecular weight (kDa) | 等电点 Isoelectric point | 跨膜结构域 Transmembrane domain | 不稳定性 系数 Instability index | 脂肪 系数 Fat index | 平均疏水性 Average hydrophobicity | 亚细胞定位预测 Subcellular localization prediction |
|---|---|---|---|---|---|---|---|---|---|---|
| AhFKBP1 | AH01G03290.1 | 648 | 215 | 24.00 | 8.44 | 0 | 52.61 | 95.63 | -0.078 | 叶绿体 |
| AhFKBP2 | AH01G12790.1 | 1872 | 623 | 70.10 | 5.33 | 0 | 42.43 | 74.00 | -0.581 | 细胞质 |
| AhFKBP3 | AH01G16560.1 | 546 | 181 | 20.45 | 7.65 | 0 | 43.44 | 94.36 | 0.155 | 叶绿体 |
| AhFKBP4 | AH02G05330.1 | 729 | 242 | 27.24 | 8.64 | 0 | 60.07 | 76.49 | -0.419 | 叶绿体 |
| AhFKBP5 | AH03G34340.1 | 705 | 234 | 25.08 | 8.10 | 0 | 27.94 | 90.00 | 0.021 | 线粒体 |
| AhFKBP6 | AH03G43820.1 | 732 | 243 | 26.30 | 8.99 | 0 | 48.70 | 81.77 | -0.256 | 叶绿体 |
| AhFKBP7 | AH03G46540.1 | 2841 | 946 | 105.08 | 5.51 | 0 | 40.18 | 91.31 | -0.160 | 细胞核 |
| AhFKBP8 | AH03G47450.1 | 633 | 210 | 22.25 | 9.24 | 0 | 45.98 | 86.81 | -0.065 | 叶绿体外膜 |
| AhFKBP9 | AH04G10810.1 | 474 | 157 | 17.82 | 7.71 | 0 | 39.81 | 88.73 | -0.292 | 细胞核 |
| AhFKBP10 | AH04G13950.1 | 708 | 235 | 25.65 | 8.26 | 1 | 57.12 | 83.40 | -0.105 | 叶绿体 |
| AhFKBP11 | AH04G16570.1 | 444 | 147 | 15.79 | 6.83 | 0 | 31.83 | 96.12 | -0.065 | 细胞外间隙 |
| AhFKBP12 | AH04G16590.1 | 1653 | 550 | 61.55 | 5.18 | 0 | 29.70 | 77.09 | -0.629 | 细胞质 |
| AhFKBP13 | AH04G16590.2 | 1653 | 550 | 61.55 | 5.18 | 0 | 29.70 | 77.09 | -0.629 | 细胞质 |
| AhFKBP14 | AH05G00870.1 | 699 | 232 | 25.26 | 8.95 | 0 | 38.62 | 78.58 | -0.241 | 叶绿体 |
| AhFKBP15 | AH05G15850.1 | 1482 | 493 | 54.74 | 5.21 | 0 | 52.79 | 57.89 | -1.059 | 细胞核 |
| AhFKBP16 | AH05G27750.1 | 381 | 126 | 14.09 | 4.83 | 0 | 28.70 | 80.32 | -0.228 | 细胞核 |
| AhFKBP17 | AH05G28070.1 | 426 | 141 | 15.58 | 7.60 | 0 | 30.39 | 76.03 | -0.383 | 细胞外间隙 |
| AhFKBP18 | AH06G10440.1 | 1065 | 354 | 40.54 | 6.18 | 1 | 43.54 | 76.07 | -0.595 | 细胞核 |
| AhFKBP19 | AH06G10440.2 | 1095 | 364 | 41.59 | 5.87 | 1 | 40.64 | 75.33 | -0.579 | 细胞核 |
| AhFKBP20 | AH06G19370.1 | 1644 | 547 | 61.66 | 5.63 | 0 | 30.55 | 76.11 | -0.588 | 细胞质 |
| AhFKBP21 | AH06G25040.1 | 507 | 168 | 18.60 | 6.92 | 0 | 29.17 | 73.04 | -0.655 | 细胞核 |
| AhFKBP22 | AH07G09740.1 | 561 | 186 | 19.89 | 6.22 | 0 | 32.84 | 76.67 | -0.439 | 细胞核 |
| AhFKBP23 | AH09G07580.1 | 1728 | 575 | 64.15 | 5.26 | 0 | 27.59 | 77.51 | -0.595 | 细胞质 |
| AhFKBP24 | AH10G05330.1 | 339 | 112 | 12.00 | 6.55 | 0 | 30.15 | 67.77 | -0.346 | 细胞外间隙 |
| AhFKBP25 | AH11G02960.1 | 570 | 189 | 20.53 | 5.02 | 0 | 21.97 | 76.30 | -0.472 | 细胞质 |
| AhFKBP26 | AH11G05460.1 | 654 | 217 | 24.20 | 8.76 | 0 | 51.33 | 95.21 | -0.093 | 叶绿体 |
| AhFKBP27 | AH11G12840.1 | 1872 | 623 | 70.19 | 5.33 | 0 | 43.67 | 73.53 | -0.594 | 细胞质 |
| AhFKBP28 | AH11G18990.1 | 780 | 259 | 28.90 | 8.52 | 0 | 37.97 | 72.01 | -0.623 | 细胞质 |
| AhFKBP29 | AH12G06190.1 | 729 | 242 | 27.22 | 8.43 | 0 | 59.13 | 76.49 | -0.417 | 叶绿体 |
| AhFKBP30 | AH12G35990.1 | 531 | 176 | 18.64 | 7.74 | 0 | 41.96 | 88.52 | -0.103 | 细胞核 |
| AhFKBP31 | AH13G06390.1 | 744 | 247 | 26.26 | 6.38 | 0 | 46.54 | 89.96 | -0.028 | 细胞质 |
| AhFKBP32 | AH13G19950.1 | 354 | 117 | 12.99 | 4.92 | 0 | 33.95 | 111.62 | 0.279 | 细胞外间隙 |
| AhFKBP33 | AH13G20760.1 | 1158 | 385 | 43.23 | 7.51 | 0 | 36.78 | 88.70 | -0.412 | 细胞质 |
| AhFKBP34 | AH13G38140.1 | 627 | 208 | 22.18 | 8.78 | 0 | 28.32 | 82.07 | -0.078 | 线粒体 |
| AhFKBP35 | AH13G38140.2 | 627 | 208 | 22.18 | 8.78 | 0 | 28.32 | 82.07 | -0.078 | 线粒体 |
| AhFKBP36 | AH13G46550.1 | 732 | 243 | 26.26 | 8.65 | 0 | 50.28 | 80.58 | -0.246 | 叶绿体 |
| AhFKBP37 | AH13G50150.1 | 636 | 211 | 22.41 | 9.23 | 0 | 44.84 | 86.40 | -0.033 | 叶绿体 |
| AhFKBP38 | AH13G52010.1 | 1731 | 576 | 64.43 | 5.20 | 0 | 30.94 | 74.79 | -0.653 | 细胞质 |
| AhFKBP39 | AH14G15180.1 | 663 | 220 | 24.47 | 5.54 | 0 | 19.82 | 76.68 | -0.564 | 细胞质 |
| AhFKBP40 | AH14G15690.1 | 1731 | 576 | 64.53 | 5.28 | 0 | 30.89 | 76.48 | -0.647 | 细胞质 |
| AhFKBP41 | AH14G15690.2 | 1455 | 484 | 53.88 | 5.20 | 0 | 26.92 | 73.26 | -0.651 | 细胞质 |
| AhFKBP42 | AH14G19580.1 | 747 | 248 | 27.34 | 9.08 | 1 | 57.84 | 84.11 | -0.121 | 叶绿体 |
| AhFKBP43 | AH14G39590.1 | 639 | 212 | 23.51 | 8.41 | 0 | 42.47 | 83.21 | -0.267 | 细胞外间隙 |
| AhFKBP44 | AH14G39590.2 | 696 | 231 | 25.47 | 7.55 | 0 | 42.93 | 81.47 | -0.216 | 细胞外间隙 |
| AhFKBP45 | AH15G07050.1 | 1485 | 494 | 54.82 | 5.25 | 0 | 50.36 | 58.56 | -1.049 | 细胞核 |
| AhFKBP46 | AH15G26530.1 | 963 | 320 | 34.89 | 5.67 | 0 | 42.65 | 86.22 | -0.093 | 细胞器膜 |
| AhFKBP47 | AH16G16120.1 | 1065 | 354 | 40.52 | 7.03 | 1 | 44.10 | 77.43 | -0.587 | 细胞核 |
| AhFKBP48 | AH16G16120.2 | 783 | 260 | 29.49 | 5.04 | 0 | 37.19 | 73.50 | -0.466 | 细胞质 |
| AhFKBP49 | AH16G24090.1 | 1644 | 547 | 61.73 | 5.41 | 0 | 29.80 | 77.17 | -0.575 | 细胞质 |
| AhFKBP50 | AH16G28470.1 | 759 | 252 | 27.68 | 8.45 | 0 | 35.83 | 81.59 | -0.354 | 细胞质 |
| AhFKBP51 | AH16G39090.1 | 1698 | 565 | 62.98 | 5.04 | 0 | 30.30 | 83.65 | -0.485 | 细胞质 |
| AhFKBP52 | AH16G42920.1 | 1776 | 591 | 65.60 | 5.09 | 0 | 29.28 | 83.27 | -0.494 | 细胞质 |
| AhFKBP53 | AH17G08750.1 | 561 | 186 | 19.94 | 6.22 | 0 | 32.23 | 77.20 | -0.439 | 细胞核 |
| AhFKBP54 | AH17G15830.1 | 534 | 177 | 19.31 | 4.88 | 0 | 23.64 | 81.41 | -0.412 | 细胞质 |
| AhFKBP55 | AH19G10630.1 | 1728 | 575 | 64.03 | 5.21 | 0 | 27.19 | 77.34 | -0.598 | 细胞质 |
| AhFKBP56 | AH20G08260.1 | 339 | 112 | 12.01 | 6.55 | 0 | 30.91 | 66.88 | -0.370 | 细胞外间隙 |
2.2 5个FKBP基因系统进化树分析
为探索花生FKBP基因家族成员间进化关系,对56个花生、64个小麦、30个水稻、17个黄瓜及24个拟南芥的FKBP基因家族成员的氨基酸系列进行比对并绘制系统进化树。结果(图1)表明,FKBP基因家族共分为7个亚群,亚群Ⅰ包含5个AhFKBP,亚群Ⅱ包含6个AhFKBP,亚群Ⅲ包含6个AhFKBP,亚群Ⅳ包含9个AhFKBP,亚群Ⅴ包含5个AhFKBP,亚群Ⅵ包含12个AhFKBP,亚群Ⅶ包含12个AhFKBP,其中AhFKBP24和AhFKBP56 2个基因编码同一种蛋白。
图1
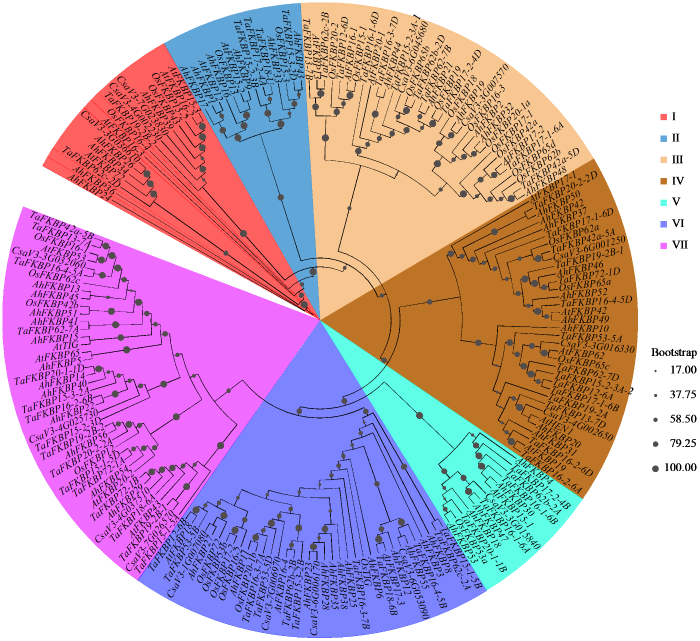
图1
花生、水稻、小麦、黄瓜、拟南芥中FKBP基因家族系统进化树
Fig.1
Phylogenetic tree of FKBP gene family in peanut, rice, wheat, cucumber and Arabidopsis
2.3 花生FKBP基因家族蛋白序列结构特征和基因结构分析
为了解AhFKBP基因家族蛋白功能区域,利用MEME软件对花生FKBP蛋白保守模块进行分析,模体宽度的最小值为6,最大值为200。如图2所示,MEME软件分析75%以上的AhFKBP蛋白含有Motif 1,近一半AhFKBP蛋白含有Motif 4,Motif 2和Motif 6也存在于多个AhFKBP蛋白中,说明这些基序在AhFKBP家族中保守性较强,推测可能是执行核心功能的关键结构域。各亚族中FKBP蛋白存在完全相同的基序模块,说明其可能具有相似功能,如AhFKBP40和AhFKBP38、AhFKBP23和AhFKBP13等。花生FKBP蛋白保守基序的数量和种类的差异反映这些蛋白结构多样性,可能存在进化上的高度分化,猜测FKBP可能在花生中具有新的功能,这需要进一步验证。
图2
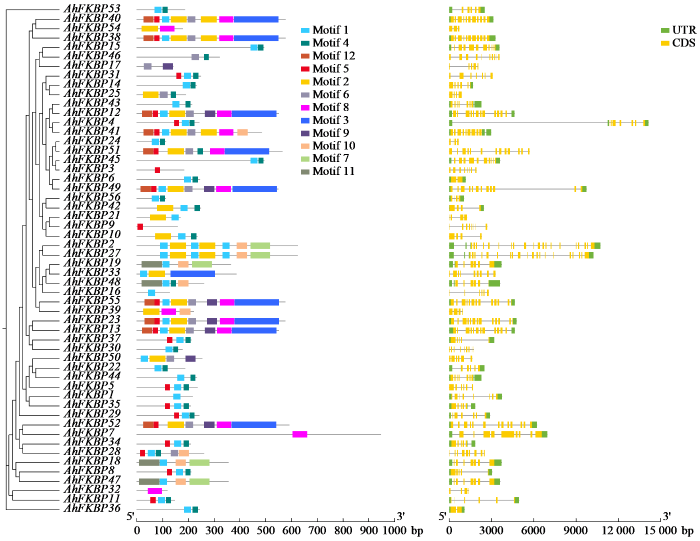
图2
AhFKBP家族保守基序和基因结构分析
Fig.2
Analysis of conserved motifs and gene structures in the AhFKBP family
基因结构分析表明,所有AhFKBP均含有多个CDS区,所有AhFKBP都具有内含子,AhFKBP41的内含子最长,基因的CDS区越多内含子就越多,AhFKBP2等内含子长短不一,基因结构较复杂。19个AhFKBP不含3’UTR和5’UTR,除这些外其余均含有3’UTR,但有1个不含5’UTR。综上,同一亚族基因结构和保守基序相似,如AhFKBP2和AhFKBP27、AhFKBP47和AhFKBP18,表明同一亚家族基因可能具有相似的功能。
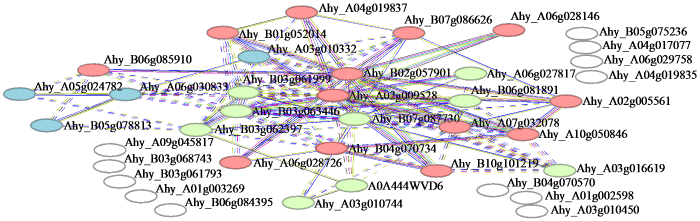
通过STRING在线预测AhFKBP蛋白间相互作用,蛋白间相互作用越强,连接的线段越多。由图3可知,可能具有相互作用的蛋白可分为3类(以小球颜色区分),第1类中的13个蛋白具有相互作用,第2类有9个蛋白,第3类有4个蛋白。AhFKBP29(Ahy-B02g057901)是网络核心蛋白,与多个蛋白具有相互作用,推测其可能在细胞生理过程扮演重要角色,应重点研究。
图3
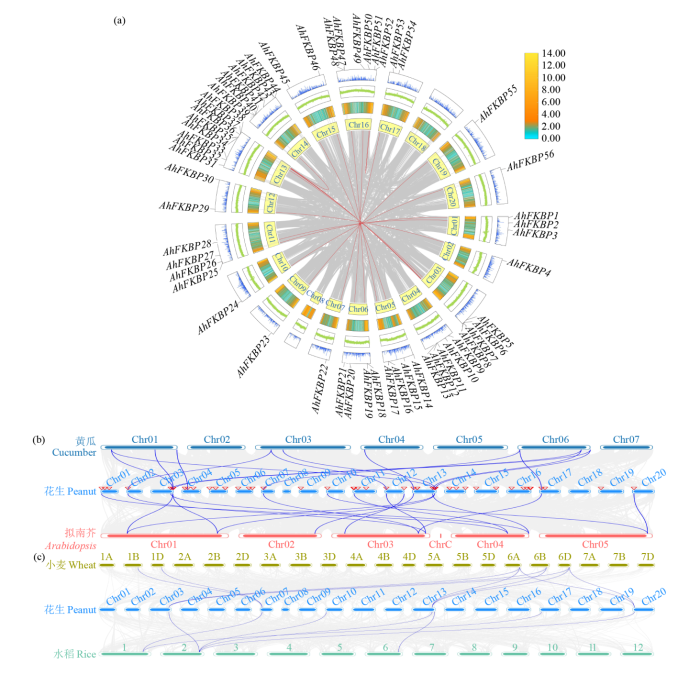
2.4 AhFKBP基因家族染色体定位和共线性分析
如图4所示,AhFKBP不均匀分布在20条染色体,其中以染色体13、14和16分布较多。图4中蓝色连接线段表示基因出现片段重复事件,可知AhFKBP51和AhFKBP52为串联重复,位于染色体16,两者不在同一亚家族中,不同染色体间共出现15次染色体间片段重复事件,可能发生基因扩张,染色体7和12出现多次重复事件,是基因组重组活跃区域,推测AhFKBP在这些染色体上富集。进一步分析花生FKBP基因家族成员间、花生FKBP与拟南芥、小麦、水稻、黄瓜FKBP基因间的共线性(图5a),发现花生FKBP基因家族成员间存在14个共线性基因(图5b),与拟南芥共有16对共线基因,与黄瓜有14对共线基因,这些共线基因集中在花生的10条染色体上。花生与小麦和水稻的共线基因分别为10和7个,较集中的分布在5条染色体上(图5c),这说明花生与拟南芥和黄瓜亲缘关系较近。
图4
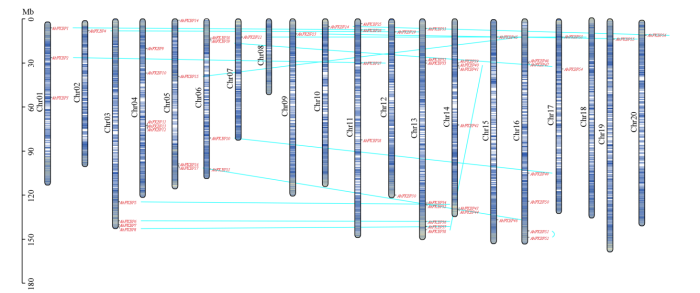
图5
2.5 花生FKBP基因家族顺式作用元件分析
为探索调控FKBP基因表达及其可能参与的信号通路,利用Plant CARE分析花生FKBP基因翻译起始位点(ATG)上游2000 bp非编码序列的顺式作用元件。由图6可知,鉴定出的顺式作用元件大致可分为4个MeJA反应性元件,包括MeJA反应性元件(TGACG基序,CGTCA基序)、脱落酸反应元件ABRE和赤霉素反应元件P-box和GARE基序;胁迫响应因子如厌氧诱导ARE、低温反应元件LTR、STRE及光响应元件G-box。花生FKBP基因家族成员启动子大多存在胁迫响应因子ARE,占预测总顺式作用元件数量的20.5%,其次是STRE,占比14.8%,胁迫响应元件占比35%以上,这增强了AhFKBP响应胁迫的可能性。结合AhFKBP染色体定位结果发现,不同染色体上片段重复基因具有相似的表达元件,如AhFKBP5和AhFKBP34、AhFKBP35,这表明不同FKBP基因在花生中受到共同因子调控,表现出相似的表达模式。
图6
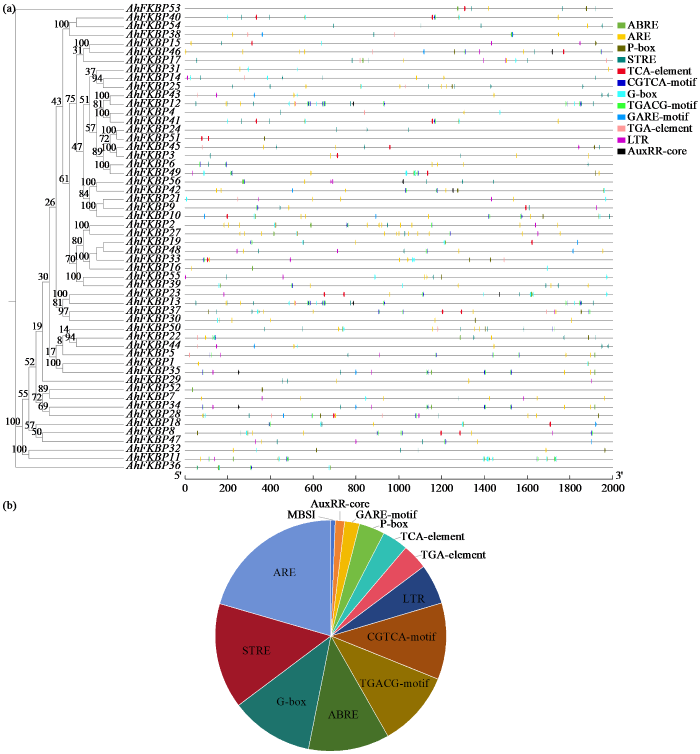
图6
花生FKBP基因启动子区顺式作用元件预测和量化分析
Fig.6
Prediction and quantitative analysis of cis-acting elements in promoter region of peanut FKBP gene
2.6 花生FKBP基因组的表达分析
图7
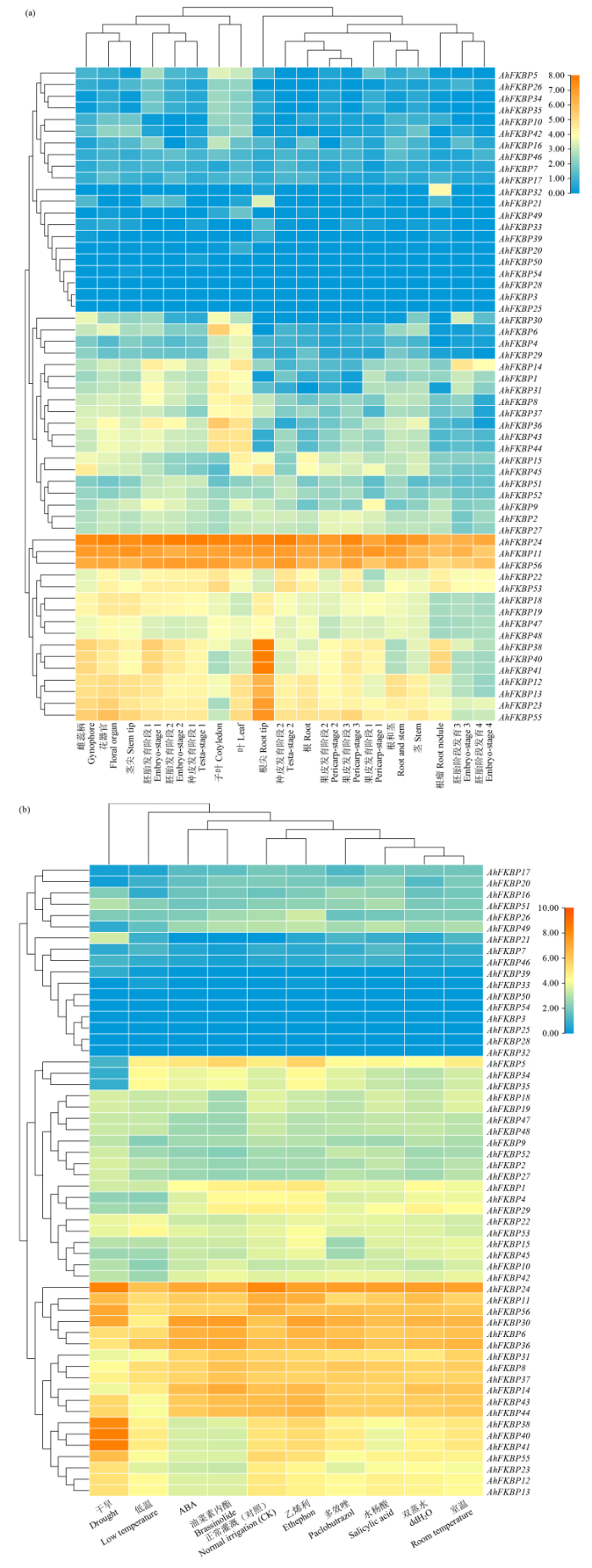
图7
FKBP基因家族在花生不同组织及不同处理下表达模式分析
Fig.7
Analysis of the expression patterns of the FKBP gene family in different tissues and under different treatments of peanuts
2.7 花生FKBP基因组功能注释
图8
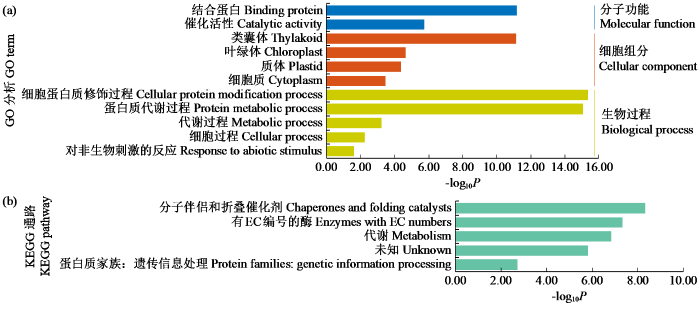
3 讨论
关于FKBP基因家族的研究主要集中于酵母和哺乳动物中,有关高等植物的研究较少,近年来,植物FKBP基因家族受到关注。研究[21-22]发现,植物FKBP基因家族编码的蛋白质主要定位于叶绿体、细胞质、细胞核等,通过与其他蛋白质结合可参与响应生物和非生物胁迫,包括病原体胁迫、干旱、低温和热胁迫等,如AtFKBP62-65-HSP90.1形成异源三聚体调节热激转录因子Hsfsa2活性,参与调控拟南芥响应热胁迫,小麦中TaBI-1与内质网膜上的TaFKBP62相互作用调控小麦响应热胁迫,水稻OsFKBP20-1a与SUMO结合酶(Sce)结合,参与水稻响应高温胁迫,OsFKBP20-1b与OsFKBP20- 1a高度同源,高温和干旱诱导该基因表达。FKBP基因家族还参与植物生长发育,拟南芥FKBP42突变体发育迟缓且根茎呈螺旋生长,FKBP72通过参与超长脂肪酸(VLCFAs)的合成影响植物生长发育,该基因功能受损时VLCFAs合成中断,细胞分裂发生紊乱,FKBP12作为植物中最小的FKBP与AtFIP37结合参与mRNA剪接、细胞周期调控和胚胎发育,也可参与调控DNA结合蛋白的核运输[23
为全面了解花生FKBP基因家族的基本特征,本研究基于花生全基因数据共鉴定出56个FKBP基因家族成员,对其染色体分布、系统发育关系、基序预测、基因结构、顺式作用元件和表达模式进行综合分析。花生FKBP基因家族成员共分布在20条染色体上,在染色体7和12发生富集,并出现了串联重复和多个片段重复事件,进一步分析基因家族成员在染色体间的共线性,发现共有14个共线性基因,这加大了AhFKBP的同源性。对家族成员编码的蛋白质进行理化性质鉴定后发现,除AhFKBP3、AhFKBP5和AhFKBP32平均疏水性大于0外,大部分蛋白为稳定的亲水性蛋白,亚细胞定位预测大部分蛋白位于细胞质、细胞核和叶绿体上,说明花生FKBP可能在信号传导、光合作用等方面产生一定影响。系统进化树分析结果表明AhFKBP基因家族分为7个亚群,目前研究较多的几个FKBP,如AtFKBP16-3参与光合作用调控,同样定位于第一亚群的AhFKBP26和AhFKBP36可能也具有类似功能,这需要进一步研究,类似的还有AtFKBP43作为分子伴侣复合物(与HSP90互作)参与热激响应和信号传导,对于定位于同一亚家族的AhFKBP且具有3个TPR结构域的AhFKBP应重点关注。
蛋白序列结构和基因结构分析可知,Motif 1、4、2和6在AhFKBP家族中保守性较强,保守基序数量和种类的差异象征着FKBP在花生中具有新的功能。种间共线性分析可知,花生FKBP与拟南芥和黄瓜FKBP分别有16个和14个同源基因,这说明花生与这2种植物亲缘关系较近。对花生FKBP家族顺式作用元件分析可知,花生FKBP基因家族启动子中含有多个顺式作用元件,包括MeJA反应性元件、脱落酸反应元件ABRE、赤霉素反应元件P-box和GARE基序、光响应元件G-box和多个胁迫响应元件ARE、LTR、TCA,表明花生FKBP基因家族可能参与花生生长发育和多种逆境响应过程。AhFKBP基因家族成员在不同组织中特异性表达,如AhFKBP38、AhFKBP40和AhFKBP41在根尖细胞中表达量较高,但在根茎组织中表达量较低,且这3个基因响应干旱胁迫,后续应对这些基因加以重视。通过以上对花生FKBP基因家族机制和功能分析,为今后更深入研究该基因家族抗逆性提供了理论基础。
4 结论
在花生基因组中共鉴定出56个AhFKBP基因家族成员,分为7个亚族,不均匀分布在20条染色体上。花生FKBP基因家族成员间及花生与拟南芥、黄瓜的FKBP具有多对共线性基因,在进化过程中表现出一定的共线模式。花生FKBP存在多个胁迫响应因子,如ARE、STRE等反应元件,表明该基因可能参与调控逆境胁迫过程。
参考文献
Abiotic stress responses in plants
DOI:10.1038/s41576-021-00413-0 [本文引用: 1]
Cyclophilins and their functions in abiotic stress and plant-microbe interactions
Plants have developed a variety of mechanisms and regulatory pathways to change their gene expression profiles in response to abiotic stress conditions and plant–microbe interactions. The plant–microbe interaction can be pathogenic or beneficial. Stress conditions, both abiotic and pathogenic, negatively affect the growth, development, yield and quality of plants, which is very important for crops. In contrast, the plant–microbe interaction could be growth-promoting. One of the proteins involved in plant response to stress conditions and plant–microbe interactions is cyclophilin. Cyclophilins (CyPs), together with FK506-binding proteins (FKBPs) and parvulins, belong to a big family of proteins with peptidyl-prolyl cis-trans isomerase activity (Enzyme Commission (EC) number 5.2.1.8). Genes coding for proteins with the CyP domain are widely expressed in all organisms examined, including bacteria, fungi, animals, and plants. Their different forms can be found in the cytoplasm, endoplasmic reticulum, nucleus, chloroplast, mitochondrion and in the phloem space. They are involved in numerous processes, such as protein folding, cellular signaling, mRNA processing, protein degradation and apoptosis. In the past few years, many new functions, and molecular mechanisms for cyclophilins have been discovered. In this review, we aim to summarize recent advances in cyclophilin research to improve our understanding of their biological functions in plant defense and symbiotic plant–microbe interactions.
Revisiting the role of plant transcription factors in the battle against abiotic stress
DOI:10.3390/ijms19061634
URL
[本文引用: 1]

Owing to diverse abiotic stresses and global climate deterioration, the agricultural production worldwide is suffering serious losses. Breeding stress-resilient crops with higher quality and yield against multiple environmental stresses via application of transgenic technologies is currently the most promising approach. Deciphering molecular principles and mining stress-associate genes that govern plant responses against abiotic stresses is one of the prerequisites to develop stress-resistant crop varieties. As molecular switches in controlling stress-responsive genes expression, transcription factors (TFs) play crucial roles in regulating various abiotic stress responses. Hence, functional analysis of TFs and their interaction partners during abiotic stresses is crucial to perceive their role in diverse signaling cascades that many researchers have continued to undertake. Here, we review current developments in understanding TFs, with particular emphasis on their functions in orchestrating plant abiotic stress responses. Further, we discuss novel molecular mechanisms of their action under abiotic stress conditions. This will provide valuable information for understanding regulatory mechanisms to engineer stress-tolerant crops.
From organ transplantation to plant biology
DOI:10.1104/pp.103.900108 URL [本文引用: 1]
Peptidylprolyl cis/trans isomerases (immunophilins): biological diversity-targets-functions
DOI:10.2174/1568026033451862 URL [本文引用: 1]
Immunophilins in protein folding and immunosuppression
Lymphocyte activation requires the transmission of signals from molecules at the plasma membrane to nuclear signals that regulate gene expression. In recent years, several immunosuppressive compounds have been used as probes to identify important and potentially novel molecules involved in lymphocyte signal transduction processes. The immunosuppressants cyclosporin A (CsA), FK506, and rapamycin have been studied in particular detail. Two distinct classes of immunosuppressant binding proteins have been identified, and collectively termed immunophilins. The cyclophilin family of immunophilins binds CsA, whereas the FK506-binding protein (FKBP) family binds FK506 and rapamycin. This review will discuss both the endogenous functions of immunophilins as well as their roles in mediating immunosuppression.
Chemistry and biology of the immunophilins and their immunosuppressive ligands
PMID:1702904
Cyclosporin A, FK506, and rapamycin are inhibitors of specific signal transduction pathways that lead to T lymphocyte activation. These immunosuppressive agents bind with high affinity to cytoplasmic receptors termed immunophilins (immunosuppressant binding proteins). Studies in this area have focused on the structural basis for the molecular recognition of immunosuppressants by immunophilins and the biological consequences of their interactions. Defining the biological roles of this emerging family of receptors and their ligands may illuminate the process of protein trafficking in cells and the mechanisms of signal transmission through the cytoplasm.
Probing immunosuppressant action with a nonnatural immunophilin ligand
PMID:1700475
The immunosuppressants FK506 and rapamycin bind to the same immunophilin, FK506 binding protein (FKBP), and inhibit distinct signal transduction pathways in T lymphocytes. A nonnatural immunophilin ligand, 506BD, which contains only the common structural elements of FK506 and rapamycin, was synthesized and found to be a high-affinity ligand of FKBP and a potent inhibitor of FKBP rotamase activity. Whereas 506BD does not interfere with T cell activation, it does block the immunosuppressive effects of both FK506 and rapamycin. Thus, the common immunophilin binding element of these immunosuppressants, which is responsible for rotamase inhibition, is fused to different effector elements, resulting in the inhibition of different signaling pathways. Inhibition of rotamase activity is an insufficient requirement for mediating these effects.
Isolation and sequence of an FK506-binding protein from N.crassa which catalyses protein folding
DOI:10.1038/346674a0
Developmental regulation of FKBP65: an ER-localized extracellular matrix binding-protein
FKBP65 (65-kDa FK506-binding protein) is a member of the highly conserved family of intracellular receptors called immunophilins. All have the property of peptidyl-prolyl cis-trans isomerization, and most have been implicated in folding and trafficking events. In an earlier study, we identified that FKBP65 associates with the extracellular matrix protein tropoelastin during its transport through the cell. In the present study, we have carried out a detailed investigation of the subcellular localization of FKBP65 and its relationship to tropoelastin. Using subcellular fractionation, Triton X-114 phase separation, protease protection assays, and immunofluorescence microscopy (IF), we have identified that FKBP65 is contained within the lumen of the endoplasmic reticulum (ER). Subsequent IF studies colocalized FKBP65 with tropoelastin and showed that the two proteins dissociate before reaching the Golgi apparatus. Immunohistochemical localization of FKBP65 in developing lung showed strong staining of vascular and airway smooth muscle cells. Similar areas stained positive for the presence of elastic fibers in the extracellular matrix. The expression of FKBP65 was investigated during development as tropoelastin is not expressed in adult tissues. Tissue-specific expression of FKBP65 was observed in 12-d old mouse tissues; however, the pattern of expression of FKBP65 was not restricted to those tissues expressing tropoelastin. This suggests that additional ligands for FKBP65 likely exist within the ER. Remarkably, in the adult tissues examined, FKBP65 expression was absent or barely detectable. Taken together, these results support an ER-localized FKBP65-tropoelastin interaction that occurs specifically during growth and development of tissues.
Immunophilins and parvulins. superfamily of peptidyl prolyl isomerases in Arabidopsis
Immunophilins are defined as receptors for immunosuppressive drugs including cyclosporin A, FK506, and rapamycin. The cyclosporin A receptors are referred to as cyclophilins (CYPs) and FK506- and rapamycin-binding proteins are abbreviated as FKBPs. These two groups of proteins (collectively called immunophilins) share little sequence homology, but both have peptidyl prolyl cis/trans isomerase (PPIase) activity that is involved in protein folding processes. Studies have identified immunophilins in all organisms examined including bacteria, fungi, animals, and plants. Nevertheless, the physiological function of immunophilins is poorly understood in any organism. In this study, we have surveyed the genes encoding immunophilins in Arabidopsis genome. A total of 52 genes have been found to encode putative immunophilins, among which 23 are putative FKBPs and 29 are putative CYPs. This is by far the largest immunophilin family identified in any organism. Both FKBPs and CYPs can be classified into single domain and multiple domain members. The single domain members contain a basic catalytic domain and some of them have signal sequences for targeting to a specific organelle. The multiple domain members contain not only the catalytic domain but also defined modules that are involved in protein-protein interaction or other functions. A striking feature of immunophilins in Arabidopsis is that a large fraction of FKBPs and CYPs are localized in the chloroplast, a possible explanation for why plants have a larger immunophilin family than animals. Parvulins represent another family of PPIases that are unrelated to immunophilins in protein sequences and drug binding properties. Three parvulin genes were found in Arabidopsis genome. The expression of many immunophilin and parvulin genes is ubiquitous except for those encoding chloroplast members that are often detected only in the green tissues. The large number of genes and diversity of structure domains and cellular localization make PPIases a versatile superfamily of proteins that clearly function in many cellular processes in plants.
Genome-wide analysis of genes encoding FK506-binding proteins in rice
DOI:10.1007/s11103-009-9547-1 URL [本文引用: 1]
Genome-wide analysis and environmental response profiling of the FK506-binding protein gene family in maize (Zea mays L.)
DOI:10.1016/j.gene.2012.01.094 URL [本文引用: 1]
Genome-wide identification and analysis of FK506-binding protein family gene family in strawberry (Fragaria×ananassa)
DOI:10.1016/j.gene.2013.08.056 URL [本文引用: 1]
Genome-wide identification and analysis of FK506-binding protein gene family in peach (Prunus persica)
DOI:10.1016/j.gene.2013.10.059
PMID:24342662
[本文引用: 1]
The FKBP protein family has prolyl isomerase activity and is related in function to cyclophilins. FKBPs are known to be involved in many biological processes including hormone signaling, plant growth, and stress responses through a chaperone or an isomerization of proline residues during protein folding. The availability of complete peach genome sequences allowed the identification of 21 FKBP genes by HMMER and BLAST analyses. Scaffold locations of these FKBP genes in the peach genome were determined and the protein domain and motif organization of peach FKBPs were analyzed. The phylogenetic relationships between peach FKBPs were also assessed. The expression profiles of peach FKBP gene results revealed that most peach FKBPs were expressed in all tissues, while a few peach FKBPs were specifically expressed in some of the tissues. This data could contribute to better understanding of the complex regulation of the peach FKBP gene family, and also provide valuable information for further research in peach functional genomics. Copyright © 2013 Elsevier B.V. All rights reserved.
大麦FKBP基因家族的鉴定与表达分析
Characterization of Arabidopsis thaliana AtFKBP42 that is membrane-bound and interacts with Hsp90
DOI:10.1046/j.1365-313X.2002.01420.x
URL
[本文引用: 1]
The twisted dwarf1 (twd1) mutant from Arabidopsis thaliana was identified in a screen for plant architecture mutants. The TWD1 gene encodes a 42 kDa FK506‐binding protein (AtFKBP42) that possesses similarity to multidomain PPIases such as mammalian FKBP51 and FKBP52, which are known to be components of mammalian steroid hormone receptor complexes. We report here for the first time the stoichiometry and dissociation constant of a protein complex from Arabidopsis that consists of AtHsp90 and AtFKBP42. Recombinant AtFKBP42 prevents aggregation of citrate synthase in almost equimolar concentrations, and can be cross‐linked to calmodulin. In comparison to one active and one inactive FKBP domain in FKBP52, AtFKBP42 lacks the PPIase active FKBP domain. While FKBP52 is found in the cytosol and translocates to the nucleus, AtFKBP42 was predicted to be membrane‐localized, as shown by electron microscopy.
A rice immunophilin homolog, OsFKBP12, is a negative regulator of both biotic and abiotic stress responses
DOI:10.3390/ijms21228791
URL
[本文引用: 1]
A class of proteins that were discovered to bind the immunosuppressant drug FK506, called FK506-binding proteins (FKBPs), are members of a sub-family of immunophilins. Although they were first identified in human, FKBPs exist in all three domains of life. In this report, a rice FKBP12 homolog was first identified as a biotic stress-related gene through suppression subtractive hybridization screening. By ectopically expressing OsFKBP12 in the heterologous model plant system, Arabidopsis thaliana, for functional characterization, OsFKBP12 was found to increase susceptibility of the plant to the pathogen, Pseudomonas syringae pv. tomato DC3000 (Pst DC3000). This negative regulatory role of FKBP12 in biotic stress responses was also demonstrated in the AtFKBP12-knockout mutant, which exhibited higher resistance towards Pst DC3000. Furthermore, this higher-plant FKBP12 homolog was also shown to be a negative regulator of salt tolerance. Using yeast two-hybrid tests, an ancient unconventional G-protein, OsYchF1, was identified as an interacting partner of OsFKBP12. OsYchF1 was previously reported as a negative regulator of both biotic and abiotic stresses. Therefore, OsFKBP12 probably also plays negative regulatory roles at the convergence of biotic and abiotic stress response pathways in higher plants.
Arabidopsis ROF1 (FKBP62) modulates thermotolerance by interacting with HSP90.1 and affecting the accumulation of HsfA2-regulated sHSPs
DOI:10.1111/tpj.2009.59.issue-3 URL [本文引用: 1]
SUMO-conjugating enzyme (Sce) and FK506-binding protein (FKBP) encoding rice (Oryza sativa L.) genes:genome-wide analysis, expression studies and evidence for their involvement in abiotic stress response
DOI:10.1007/s00438-008-0318-5
PMID:18219493
[本文引用: 1]
We report an in-depth characterization of two major stress proteins namely SUMO-conjugating enzyme (Sce) and peptidyl prolyl cis-trans isomerase (PPIase) in rice (Oryza sativa L.). Sce mediates addition of SUMO group to various cell proteins, through process referred to as SUMOylation. Rice nuclear genome has two putative genes encoding the Sce protein (OsSce1 and OsSce2). PCR-amplified full-length OsSce1 cDNA functionally complemented the growth defect in yeast cells lacking the equivalent Ubc9 protein (ScDeltaubc9). RT-PCR analysis showed that transcript levels of OsSce1 and OsSce2 in rice seedlings were regulated by temperature stress. OsSce1 protein was localized to the nucleus in onion epidermal cells as evidenced by the transient GFP expression analysis following micro-projectile gun-based shooting of an OsSce1-GFP fusion construct. PPIase proteins assist molecular chaperones in reactions associated with protein folding and protein transport across membrane. There are 23 putative genes encoding for FK506-binding proteins (FKBPs; specific class of PPIase) in rice genome. OsFKBP20 cDNA was isolated as a stress-inducible EST clone. Largest ORF of 561 bases in OsFKBP20 showed characteristic FK506-binding domain at N-terminus and a coiled-coil motif at C-terminus. RNA expression analysis indicated that OsFKBP20 transcript is heat-inducible. OsFKBP20 over-expression in yeast endowed capacity of high temperature tolerance to yeast cells. Yeast two-hybrid analysis showed that OsSce1 protein physically interacts with the OsFKBP20 protein. It is thus proposed that OsSce1 and OsFKBP20 proteins in concert mediate the stress response of rice plants.
TWISTED DWARF1, a unique plasma membraneanchored immunophilin- like protein, interacts with Arabidopsis multidrug resistance-like transporters AtPGP1 and AtPGP19
Null-mutations of the Arabidopsis FKBP-like immunophilin TWISTED DWARF1 (TWD1) gene cause a pleiotropic phenotype characterized by reduction of cell elongation and disorientated growth of all plant organs. Heterologously expressed TWD1 does not exhibit cis-trans-peptidylprolyl isomerase (PPIase) activity and does not complement yeast FKBP12 mutants, suggesting that TWD1 acts indirectly via protein-protein interaction. Yeast two-hybrid protein interaction screens with TWD1 identified cDNA sequences that encode the C-terminal domain of Arabidopsis multidrug-resistance-like ABC transporter AtPGP1. This interaction was verified in vitro. Mapping of protein interaction domains shows that AtPGP1 surprisingly binds to the N-terminus of TWD1 harboring the cis-trans peptidyl-prolyl isomerase-like domain and not to the tetratrico-peptide repeat domain, which has been shown to mediate protein-protein interaction. Unlike all other FKBPs, TWD1 is shown to be an integral membrane protein that colocalizes with its interacting partner AtPGP1 on the plasma membrane. TWD1 also interacts with AtPGP19 (AtMDR1), the closest homologue of AtPGP1. The single gene mutation twd1-1 and double atpgp1-1/atpgp19-1 (atmdr1-1) mutants exhibit similar phenotypes including epinastic growth, reduced inflorescence size, and reduced polar auxin transport, suggesting that a functional TWD1-AtPGP1/AtPGP19 complex is required for proper plant development.
Very-long-chain fatty acids are involved in polar auxin transport and developmental patterning in Arabidopsis
DOI:10.1105/tpc.109.071209
URL
Very-long-chain fatty acids (VLCFAs) are essential for many aspects of plant development and necessary for the synthesis of seed storage triacylglycerols, epicuticular waxes, and sphingolipids. Identification of the acetyl-CoA carboxylase PASTICCINO3 and the 3-hydroxy acyl-CoA dehydratase PASTICCINO2 revealed that VLCFAs are important for cell proliferation and tissue patterning. Here, we show that the immunophilin PASTICCINO1 (PAS1) is also required for VLCFA synthesis. Impairment of PAS1 function results in reduction of VLCFA levels that particularly affects the composition of sphingolipids, known to be important for cell polarity in animals. Moreover, PAS1 associates with several enzymes of the VLCFA elongase complex in the endoplasmic reticulum. The pas1 mutants are deficient in lateral root formation and are characterized by an abnormal patterning of the embryo apex, which leads to defective cotyledon organogenesis. Our data indicate that in both tissues, defective organogenesis is associated with the mistargeting of the auxin efflux carrier PIN FORMED1 in specific cells, resulting in local alteration of polar auxin distribution. Furthermore, we show that exogenous VLCFAs rescue lateral root organogenesis and polar auxin distribution, indicating their direct involvement in these processes. Based on these data, we propose that PAS1 acts as a molecular scaffold for the fatty acid elongase complex in the endoplasmic reticulum and that the resulting VLCFAs are required for polar auxin transport and tissue patterning during plant development.
Hormonal control of cell proliferation requires PASTICCINO genes
DOI:10.1104/pp.102.019026
PMID:12857804
PASTICCINO (PAS) genes are required for coordinated cell division and differentiation during plant development. In loss-of-function pas mutants, plant aerial tissues showed ectopic cell division that was specifically enhanced by cytokinins, leading to disorganized tumor-like tissue. To determine the role of the PAS genes in controlling cell proliferation, we first analyzed the expression profiles of several genes involved in cell division and meristem function. Differentiated and meristematic cells of the pas mutants were more competent for cell division as illustrated by the ectopic and enlarged expression profiles of CYCLIN-DEPENDENT KINASE A and CYCLIN B1. The expression of meristematic homeobox genes KNOTTED-LIKE IN ARABIDOPSIS (KNAT2, KNAT6), and SHOOT MERISTEMLESS was also increased in pas mutants. Moreover, the loss of meristem function caused by shoot meristemless mutation can be suppressed by pas2. The KNAT2 expression pattern defines an enlarged meristematic zone in pas mutants that can be mimicked in wild type by cytokinin treatment. Cytokinin induction of the primary cytokinin response markers, ARABIDOPSIS RESPONSE REGULATOR (ARR5 and ARR6), was enhanced and lasted longer in pas mutants, suggesting that PAS genes in wild type repress cytokinin responses. The expression of the cytokinin-regulated cyclin D, cyclin D3.1, was nonetheless not modified in pas mutants. However, primary auxin response genes were down-regulated in pas mutants, as shown by a lower auxin induction of IAA4 and IAA1 genes, demonstrating that the auxin response was also modified. Altogether, our results suggest that PAS genes are involved in the hormonal control of cell division and differentiation.
An Arabidopsis immunophilin, AtFKBP12, binds to AtFIP37 (FKBP interacting protein) in an interaction that is disrupted by FK506
DOI:10.1046/j.1365-313X.1998.00248.x
URL
AtFKBP12 is an Arabidopsis cDNA that encodes a protein similar to the mammalian immunophilin, FKBP12. AtFKBP12 was used as ‘bait’ in a yeast 2‐hybrid system to screen for cDNAs in Arabidopsis encoding proteins that bind to FKBP12. Two partial cDNAs were recovered encoding the C‐terminus of a protein we have called\n \n Arabidopsis thaliana\n \n FKBP12 interacting protein 37 (AtFIP37). AtFIP37 is similar to a mammalian protein, FAP48, that also binds to FKBP12. The interaction between AtFKBP12 and AtFIP37 in the 2‐hybrid system, as assessed by histidine auxotrophy and β‐galactosidase activity, was disrupted by FK506, but not by cyclosporin A, a drug that binds to cyclophilin A. AtFIP37 was also shown to bind\n \n in vitro\n \n to AtFKBP12 in GST‐fusion protein binding assays. The binding was abolished by prior incubation of AtFKBP12 with FK506. These findings indicate that an Arabidopsis FKBP12 ortholog encodes a protein that binds FK506 and that the interaction between AtFKBP12 and AtFIP37 may involve the FK506 binding site of AtFKBP12. The interaction provides interesting new opportunities for controlling protein:protein interactions\n \n in vivo\n \n in plants.
PwHAP5, a CCAAT-binding transcription factor, interacts with PwFKBP12 and plays a role in pollen tube growth orientation in Picea wilsonii
DOI:10.1093/jxb/err120
PMID:21784992
[本文引用: 1]
The HAP complex occurs in many eukaryotic organisms and is involved in multiple physiological processes. Here it was found that in Picea wilsonii, HAP5 (PwHAP5), a putative CCAAT-binding transcription factor gene, is involved in pollen tube development and control of tube orientation. Quantitative real-time reverse transcription-PCR showed that PwHAP5 transcripts were expressed strongly in germinating pollen and could be induced by Ca(2+). Overexpression of PwHAP5 in pollen altered pollen tube orientation, whereas the tube with PwHAP5RNAi showed normal growth without diminishing pollen tube growth. Furthermore, PwFKBP12, which encodes an FK506-binding protein (FKBP) was screened and a bimolecular fluorescence complementation assay performed to confirm the interaction of PwHAP5 and PwFKBP12 in vivo. Transient expression of PwFKBP12 in pollen showed normal pollen tube growth, whereas the tube with PwFKBP12RNAi bent. The phenotype of overexpression of HAP5 on pollen tube was restored by FKBP12. Altogether, our study supported the role of HAP5 in pollen tube development and orientation regulation and identified FKBP12 as a novel partner to interact with HAP5 involved in the process.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}