


水稻种子在低氧状态下,主要采取2种响应策略:“静止”和“伸长”。“静止”策略通过抑制胚芽鞘的生长,保存能量以维持种子正常生命活动;“伸长”策略则是通过促进胚芽鞘快速伸长,突破水层,使萌发种子迅速脱离低氧环境,以达到有氧环境[11]。“静止”策略更加适用于短期淹水且退水快的逆境,“伸长”的耐淹策略则更适合应用于田间生产[11-12]。因此,直播栽培过程中,选择胚芽鞘伸长能力强且伸长速度快的品种,并在直播后保持适宜的水面高度,既可以保证直播稻的齐苗,又能防止杂草的生长,减少除草剂对环境的污染[13-14]。国内外学者多采用水层隔绝空气营造低氧环境,以5或10 cm淹水深度下的胚芽鞘长度为指标进行耐低氧萌发鉴定[15-
水稻在低氧条件下会发生一系列激素和酶活的变化。缺氧状态下,赤霉素3(gibberellic acid-3,GA3)和脱落酸(abscisic acid,ABA)的含量会升高,生长素(indole-3-acetic acid,IAA)的含量也会增加[19-20]。α-淀粉酶(α-amylase,α-AMY)的活性与胚芽鞘的伸长之间存在正相关性,影响胚芽鞘的生长速度[21]。钱海丰等[21]研究表明,缺氧状态下,GA3会大量合成,从而促进α-AMY的合成,促使淀粉水解,进而促进种子的萌发。在种子的萌发过程中,活性氧(reactive oxygen species,ROS)主要通过促进种子合成水解酶从而刺激种子的萌发[22-23]。在低氧条件下,种子在淹水初期会诱使乙醇脱氢酶基因表达,耐淹品种表现出较高的乙醇脱氢酶(alcohol dehydrogenase,ADH)活性[24]。
综上所述,前人对水稻耐低氧萌发种质的筛选以及耐低氧萌发品种的生理特性做了大量研究,但影响水稻耐低氧萌发能力以及导致籼稻和粳稻耐低氧萌发能力差异的关键生理指标还不明确。为初步探究这一关键问题,本研究采用5.0和10.0 cm淹水条件,首先以胚芽鞘长度为主要指标鉴定和筛选耐低氧萌发种质资源,然后进一步测定耐低氧萌发材料相关酶活性和内源激素含量,探究耐低氧萌发种质间以及籼粳稻间耐低氧能力差异的关键生理指标,为培育耐淹萌发能力强的水稻新品种提供重要理论依据和参考。
1 材料与方法
1.1 试验材料
供试材料为收集的来自世界各地的213份粳稻和219份籼稻,共432份水稻种质资源。为打破种子休眠,减少种子休眠带来的影响,所有供试材料均置于50 ℃下处理72 h。
1.2 试验方法
每个供试材料挑选健康、饱满和均一的种子100粒,3次重复,在75%的酒精中消毒5 min,再用2% NaClO溶液灭菌15 min,最后用纯水冲洗5次。将供试材料分别放于铺有双层滤纸的培养皿(直径9 cm)中浸种48 h,每个重复挑选25粒露白的种子放于50 mL离心管中(直径2.7 cm,高度11.5 cm),设置0.5、5.0和10.0 cm 3个淹水深度,其中0.5 cm为对照处理(CK)。分别向离心管中注入纯水至0.5、5.0和10.0 cm,每天注水以保持相应淹水处理高度不变。在26±2 ℃、相对湿度75%左右的培养箱中,黑暗培养7 d。统计种子存活率(胚芽鞘继续伸长的种子数量占全部种子数量的比率),随机取10粒种子测量胚芽鞘长度,并计算耐淹指数(淹水处理下胚芽鞘长度与对照处理下胚芽鞘长度的差值/CK处理下胚芽鞘长度)。
1.3 测定指标与方法
黑暗培养7 d后,取胚芽鞘鲜样,采用源桔生物的ELISA试剂盒(中国,上海;
1.4 数据处理
使用Microsoft Excel 2021软件进行数据处理,使用SPSS 26.0进行检验方差分析和多重比较。使用GraphPad Prism 8.3.0作图。
2 结果与分析
2.1 耐低氧萌发种质资源筛选
5.0和10.0 cm淹水处理下,种子存活率和胚芽鞘长度呈极显著正相关(r=0.75,P<0.0001)。因此,本研究以萌发7 d的胚芽鞘长度为主要指标对432份水稻种质资源进行耐低氧萌发筛选,结果见网络增强出版附加材料附表1。0.5、5.0和10.0 cm淹水处理下,种子存活率的变异系数分别为44.88%、97.93%和74.83%,胚芽鞘长度分别为28.65%、63.99%和46.28%(图1和表1)。在0.5 cm处理下,种子存活率的变幅为13.33%~100.00%,胚芽鞘长度的变幅为0.55~1.93 cm;在5.0 cm淹水处理下,种子存活率的变幅为0.00%~90.00%,胚芽鞘长度的变幅为0.00~3.35 cm;在10.0 cm淹水处理下,种子存活率的变幅为0.00%~96.67%,胚芽鞘长度的变幅为0.00~3.53 cm。以上结果表明,432份水稻种质资源的种子存活率和胚芽鞘长度均表现出广泛的变异,在耐低氧萌发性上具有广泛的遗传多样性,且淹水处理会增大品种间种子存活率和胚芽鞘长度的变异。
图1
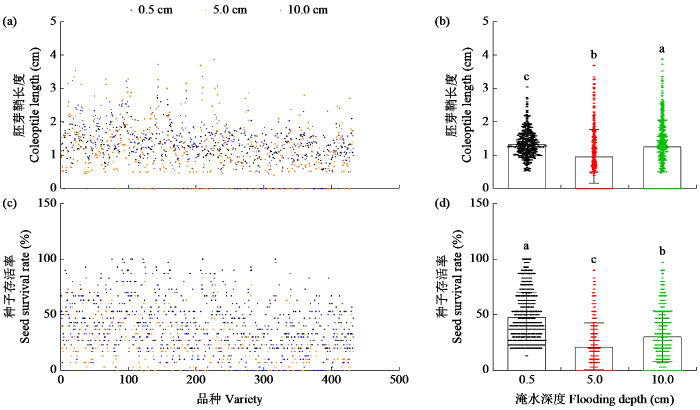
图1
不同淹水深度下水稻种质资源耐淹指标
(a)~(b) 不同淹水深度下水稻种质资源的胚芽鞘长度;(c)~(d) 不同淹水深度下水稻种质资源的存活率。不同小写字母表示差异显著(P < 0.05)。下同。
Fig.1
Submergence tolerance index of rice germplasm resources under different flooding depths
(a)~(b) The coleoptile length of rice germplasm resources under different water depths; (c)~(d) The survival rate of rice germplasm resources under different flooding depths. Different lowercase letters indicate significant differences (P < 0.05). The same below.
表1 不同淹水处理下水稻种子存活率和胚芽鞘长度的均值和变异系数
Table 1
| 淹水深度 Flooding depth (cm) | 种子存活率Seed survival rate (%) | 胚芽鞘长度Coleoptile length (cm) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 变幅 Range | 平均值 Mean | 中位数 Median | 变异系数 CV (%) | 变幅 Range | 平均值 Mean | 中位数 Median | 变异系数 CV (%) | ||
| 0.5 | 13.33~100.00 | 48.40 | 43.33 | 44.88 | 0.55~1.93 | 1.37 | 1.30 | 28.65 | |
| 5.0 | 0.00~90.00 | 21.49 | 17.67 | 97.93 | 0.00~3.35 | 1.26 | 0.85 | 63.99 | |
| 10.0 | 0.00~96.67 | 31.57 | 27.67 | 74.83 | 0.00~3.53 | 1.66 | 1.28 | 46.28 | |
表2 前10份耐低氧萌发能力强的种质资源
Table 2
| 编号 Number | 品种 Variety | 淹水深度 Flooding depth (cm) | 胚芽鞘长度 Coleoptile length (cm) | 耐淹指数 Waterlogging tolerance index | 编号 Number | 品种 Variety | 淹水深度 Flooding depth (cm) | 胚芽鞘长度 Coleoptile length (cm) | 耐淹指数 Waterlogging tolerance index |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 须谷糯 | 0.5 | 1.55 | 6 | 葡萄黄 | 0.5 | 1.36 | ||
| 5.0 | 1.92 | 0.24 | 5.0 | 2.23 | 0.64 | ||||
| 10.0 | 2.23 | 0.44 | 10.0 | 2.90 | 1.13 | ||||
| 2 | 金枝糯 | 0.5 | 1.97 | 7 | Jan-76 | 0.5 | 1.87 | ||
| 5.0 | 2.18 | 0.11 | 5.0 | 2.14 | 0.14 | ||||
| 10.0 | 2.58 | 0.31 | 10.0 | 2.72 | 0.45 | ||||
| 3 | 饿死牛 | 0.5 | 1.47 | 8 | 农垦58 | 0.5 | 1.89 | ||
| 5.0 | 1.66 | 0.13 | 5.0 | 2.03 | 0.07 | ||||
| 10.0 | 2.73 | 0.86 | 10.0 | 2.59 | 0.37 | ||||
| 4 | 闷加丁2 | 0.5 | 1.80 | 9 | 黄皮糯 | 0.5 | 1.45 | ||
| 5.0 | 2.37 | 0.32 | 5.0 | 2.96 | 1.04 | ||||
| 10.0 | 2.56 | 0.42 | 10.0 | 3.25 | 1.24 | ||||
| 5 | 博B | 0.5 | 1.37 | 10 | 辽粳287 | 0.5 | 3.03 | ||
| 5.0 | 1.54 | 0.12 | 5.0 | 3.69 | 0.22 | ||||
| 10.0 | 2.31 | 0.69 | 10.0 | 4.36 | 0.44 |
2.2 耐低氧萌发材料的表型分析
通过对432份种质资源进行筛选鉴定,粳稻品种葡萄黄和黄皮糯,籼稻品种饿死牛和博B在淹水胁迫下表现出较强的耐低氧萌发能力,为了验证该表型,进一步对4个品种进行耐低氧鉴定。对照(0.5 cm)处理下,4个材料的胚芽鞘长度无显著差异,5.0和10.0 cm的淹水处理下,4个材料的胚芽鞘长度都显著高于对照处理,且10.0 cm的淹水处理显著高于5.0 cm淹水处理(图2)。进一步分析发现,4个材料的胚芽鞘长度在对照处理下无显著差异,而在5.0和10.0 cm的淹水处理下葡萄黄和黄皮糯的胚芽鞘长度显著高于饿死牛和博B,特别是在10.0 cm淹水处理下胚芽鞘长度表现出更显著的差异(表3)。以上结果表明,粳稻品种比籼稻品种表现出更强的耐低氧萌发能力。
图2
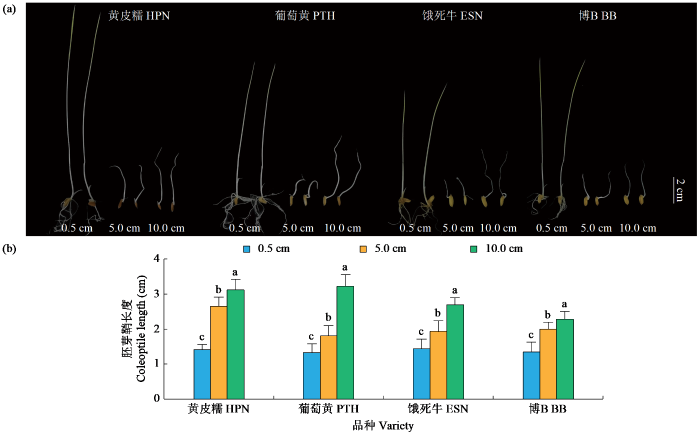
图2
黄皮糯、葡萄黄、饿死牛和博B耐低氧萌发表现
Fig.2
Germinability of Huangpinuo (HPN), Putaohuang (PTH), Esiniu (ESN) and BoB (BB) under submerged condition
表3 淹水条件下4个品种的平均胚芽鞘长度
Table 3
| 淹水深度 Flooding depth (cm) | 品种 Variety | 最小值 Min. (cm) | 最大值 Max. (cm) | 平均值 Mean (cm) | 标准 差 SD | 变异 系数 CV (%) |
|---|---|---|---|---|---|---|
| 0.5 | 饿死牛 | 1.42 | 1.52 | 1.42gh | 0.24 | 0.16 |
| 博B | 1.34 | 1.44 | 1.35ij | 0.25 | 0.19 | |
| 黄皮糯 | 1.38 | 1.58 | 1.44hi | 0.11 | 0.08 | |
| 葡萄黄 | 1.36 | 1.43 | 1.38j | 0.21 | 0.16 | |
| 5.0 | 饿死牛 | 1.89 | 2.16 | 1.92ef | 0.28 | 0.14 |
| 博B | 1.96 | 2.23 | 2.13de | 0.16 | 0.08 | |
| 黄皮糯 | 2.59 | 2.82 | 2.70bc | 0.23 | 0.09 | |
| 葡萄黄 | 1.80 | 1.93 | 1.84fg | 0.27 | 0.15 | |
| 10.0 | 饿死牛 | 2.64 | 2.87 | 2.83b | 0.16 | 0.06 |
| 博B | 2.43 | 2.43 | 2.34cd | 0.19 | 0.08 | |
| 黄皮糯 | 3.15 | 3.31 | 3.12a | 0.27 | 0.09 | |
| 葡萄黄 | 3.24 | 3.41 | 3.30a | 0.30 | 0.09 |
不同小写字母表示处理间差异显著(P < 0.05),下同。
Different lowercase letters indicate significant difference (P < 0.05), the same below.
2.3 低氧胁迫下耐低氧萌发材料的酶活性变化
在5.0 cm淹水处理下,葡萄黄的ADH活性,黄皮糯的α-AMY和ADH活性,饿死牛的α-AMY、ADH活性和ROS含量,博B的ADH活性显著高于0.5 cm处理;而葡萄黄的α-AMY活性和ROS含量,黄皮糯的ROS含量,博B的α-AMY活性和ROS含量与0.5 cm处理无显著差异(图3)。在10.0 cm淹水处理下,除葡萄黄的ROS含量与0.5 cm处理无显著差异外,4个品种的α-AMY和ADH活性都显著高于0.5 cm处理(图3)。在0.5、5.0和10.0 cm淹水处理下,粳稻品种的α-AMY、ADH活性和ROS含量都显著高于籼稻品种(表4)。以上结果表明,α-AMY、ADH活性和ROS含量在水稻耐低氧萌发中发挥重要作用,也是导致籼、粳稻耐低氧萌发能力差异的重要指标。
图3
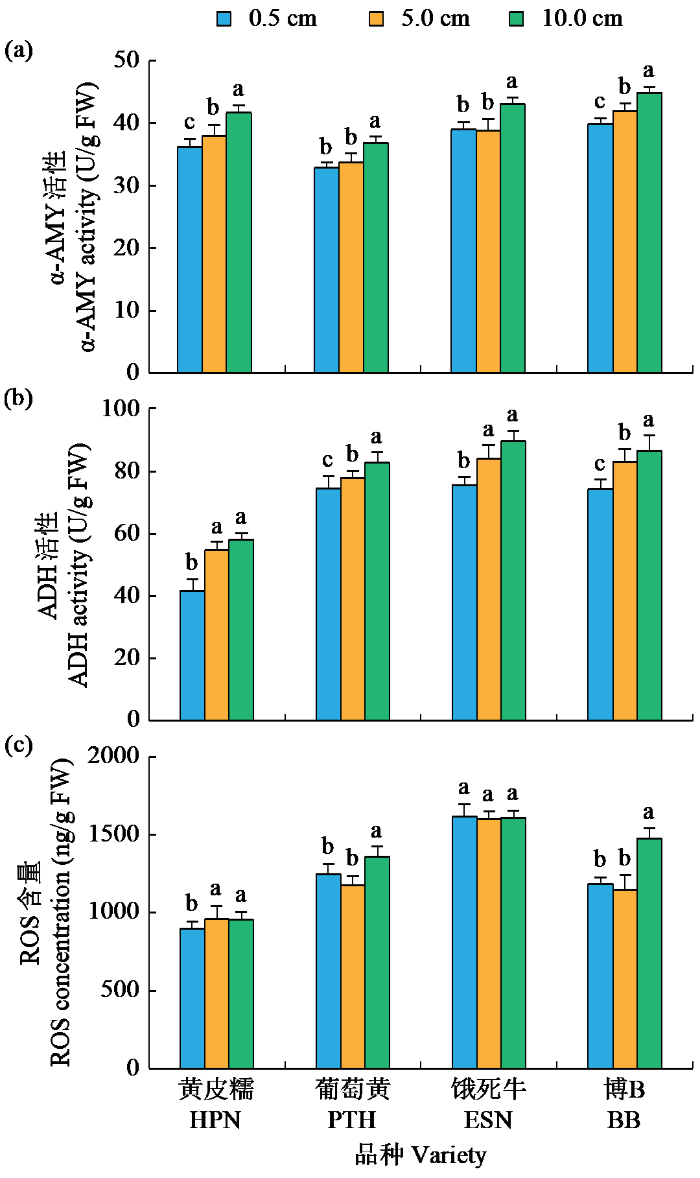
图3
不同淹水深度下耐低氧萌发材料酶活性和活性氧分析
Fig.3
Analysis of enzyme activity and ROS of hypoxia-tolerant germination materials under different flooding depths
表4 不同淹水深度下籼、粳稻相关酶活性和活性氧特征
Table 4
| 淹水深度 Flooding depth (cm) | 水稻 Rice | 项目 Item | α-AMY (U/g FW) | ADH (U/g FW) | ROS (ng/g FW) |
|---|---|---|---|---|---|
| 0.5 | 籼稻 | 最大值 | 37.60 | 77.57 | 1322.95 |
| 最小值 | 32.91 | 39.82 | 876.41 | ||
| 平均值 | 34.89c | 58.90d | 1087.69d | ||
| 标准差 | 2.88 | 16.64 | 177.10 | ||
| 变异系数 (%) | 0.08 | 0.28 | 0.16 | ||
| 粳稻 | 最大值 | 40.69 | 78.22 | 1692.42 | |
| 最小值 | 38.97 | 73.12 | 1179.65 | ||
| 平均值 | 39.88b | 75.58c | 1415.90bc | ||
| 标准差 | 0.68 | 1.83 | 220.10 | ||
| 变异系数 (%) | 0.02 | 0.02 | 0.16 | ||
| 5.0 | 籼稻 | 最大值 | 39.68 | 80.03 | 1239.08 |
| 最小值 | 33.31 | 53.46 | 920.04 | ||
| 平均值 | 36.23c | 67.09d | 1084.94cd | ||
| 标准差 | 2.34 | 11.70 | 117.09 | ||
| 变异系数 (%) | 0.06 | 0.17 | 0.11 | ||
| 粳稻 | 最大值 | 43.26 | 87.77 | 1646.52 | |
| 最小值 | 37.77 | 80.85 | 1100.81 | ||
| 平均值 | 40.85b | 84.31b | 1388.54b | ||
| 标准差 | 1.78 | 2.88 | 231.78 | ||
| 变异系数 (%) | 0.04 | 0.03 | 0.17 | ||
| 10.0 | 籼稻 | 最大值 | 42.97 | 85.20 | 1427.78 |
| 最小值 | 36.71 | 57.43 | 945.54 | ||
| 平均值 | 39.67bc | 71.23c | 1171.51bc | ||
| 标准差 | 2.47 | 12.42 | 204.58 | ||
| 变异系数 (%) | 0.06 | 0.17 | 0.17 | ||
| 粳稻 | 最大值 | 45.70 | 92.38 | 1652.19 | |
| 最小值 | 42.65 | 83.41 | 1446.48 | ||
| 平均值 | 44.37a | 88.82a | 1560.67a | ||
| 标准差 | 1.06 | 3.12 | 73.84 | ||
| 变异系数 (%) | 0.02 | 0.04 | 0.05 |
2.4 低氧胁迫下耐低氧萌发材料内源激素含量的变化
在5.0 cm淹水处理下,葡萄黄的GA3、ABA和IAA含量,黄皮糯的GA3和IAA含量,饿死牛的GA3和IAA含量,博B的IAA含量显著高于0.5 cm处理;而黄皮糯的ABA含量,饿死牛的ABA含量,博B的GA3和ABA含量与0.5 cm处理无显著差异(图4)。在10.0 cm淹水处理下,4个品种的GA3、ABA和IAA含量都显著高于0.5 cm处理(图4)。不同淹水条件下粳稻和籼稻间内源激素差异分析表明,在0.5、5.0和10.0 cm淹水处理下,粳稻品种的GA3、IAA和ABA含量均显著高于籼稻品种(表5)。以上结果表明,GA3、IAA和ABA含量在水稻耐低氧萌发以及籼粳稻耐低氧萌发能力差异中同样发挥重要作用。
图4
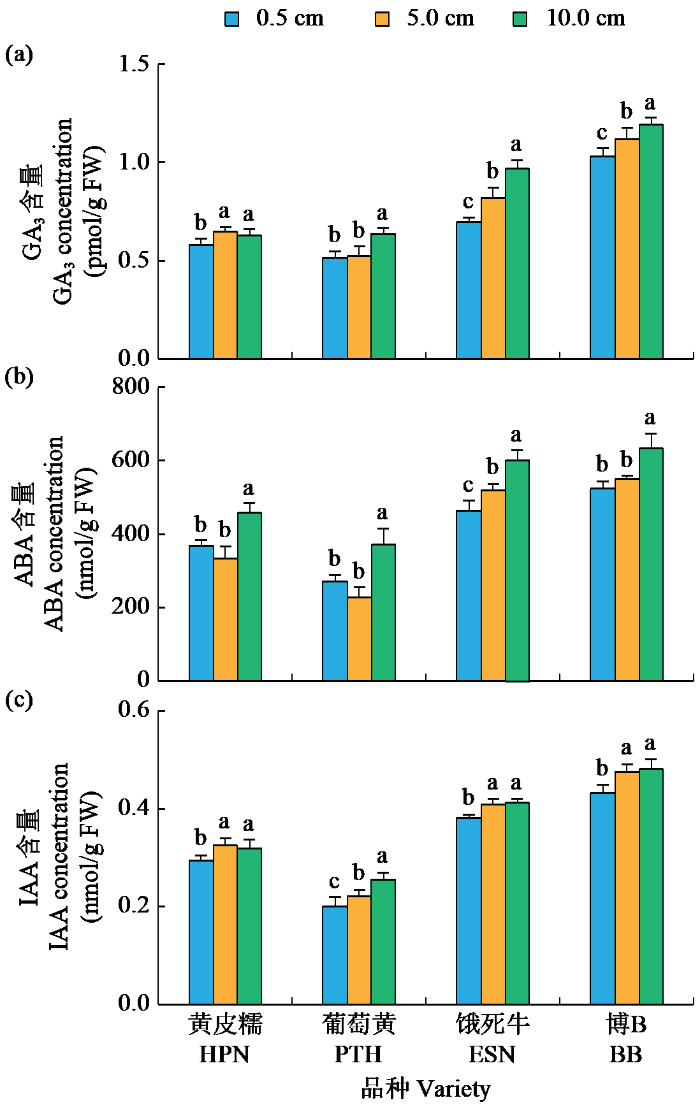
图4
不同淹水深度下耐低氧萌发材料内源激素特征
Fig.4
Endogenous auxin characteristics of hypoxia-tolerant germination materials under different flooding depths
表5 不同淹水深度下籼粳稻相关激素含量特征
Table 5
| 淹水深度 Flooding depth (cm) | 水稻 Rice | 项目 Item | GA3 (pmol/g FW) | ABA (nmol/g FW) | IAA (nmol/g FW) |
|---|---|---|---|---|---|
| 0.5 | 籼稻 | 最大值 | 0.62 | 379.86 | 0.31 |
| 最小值 | 0.50 | 266.61 | 0.19 | ||
| 平均值 | 0.56e | 326.34de | 0.25d | ||
| 标准差 | 17.32 | 48.93 | 0.05 | ||
| 变异系数 | 31.02 | 0.15 | 0.19 | ||
| 粳稻 | 最大值 | 1.07 | 545.33 | 0.45 | |
| 最小值 | 0.69 | 444.07 | 0.38 | ||
| 平均值 | 0.88 | 500.21 | 0.41 | ||
| 标准差 | 0.17c | 34.15c | 0.026b | ||
| 变异系数 | 0.19 | 0.07 | 0.06 | ||
| 5.0 | 籼稻 | 最大值 | 0.67 | 356.41 | 0.34 |
| 最小值 | 0.50 | 216.52 | 0.22 | ||
| 平均值 | 0.60d | 286.82e | 0.28c | ||
| 标准差 | 0.06 | 57.86 | 0.05 | ||
| 变异系数 | 0.11 | 0.20 | 0.19 | ||
| 粳稻 | 最大值 | 1.18 | 557.85 | 0.49 | |
| 最小值 | 0.79 | 510.69 | 0.41 | ||
| 平均值 | 0.98b | 540.30b | 0.45a | ||
| 标准差 | 0.15 | 17.32 | 0.03 | ||
| 变异系数 | 0.16 | 0.03 | 0.08 | ||
| 10.0 | 籼稻 | 最大值 | 0.67 | 482.44 | 0.34 |
| 最小值 | 0.62 | 344.42 | 0.25 | ||
| 平均值 | 0.64d | 420.80d | 0.29c | ||
| 标准差 | 0.02 | 48.60 | 0.03 | ||
| 变异系数 | 0.03 | 0.12 | 0.12 | ||
| 粳稻 | 最大值 | 1.23 | 668.43 | 0.50 | |
| 最小值 | 0.94 | 595.42 | 0.41 | ||
| 平均值 | 1.09a | 624.95a | 0.45a | ||
| 标准差 | 0.12 | 27.50 | 0.04 | ||
| 变异系数 | 0.11 | 0.04 | 0.08 |
2.5 低氧胁迫下耐低氧萌发指标间相关性分析
在5.0 cm淹水处理下,ADH与ABA、GA3、IAA、ROS、α-AMY,ABA与IAA、ROS、α-AMY,GA3与IAA、α-AMY,IAA与α-AMY有较好的正相关性(r>0.6);在10.0 cm淹水处理下,ADH与GA3、IAA、ROS,ABA与ROS,GA3与IAA、ROS、α-AMY,IAA与ROS、α-AMY呈正相关(r>0.6)(表6)。而在5.0和10.0 cm淹水处理下,胚芽鞘长度与α-AMY、GA3、IAA呈正相关(r>0.9)。综上,α-AMY活性、GA3和IAA含量在水稻耐低氧萌发中发挥更加重要的作用,同时也是导致粳稻和籼稻耐低氧萌发能力差异的主要因素。
表6 不同淹水处理下生理指标间及与胚芽鞘长度间的相关性分析
Table 6
| 指标 Index | 淹水深度 Flooding depth (cm) | 胚芽鞘长度 Coleoptile length | α-AMY活性 α-AMY activity | ADH活性 ADH activity | ROS含量 ROS content | GA3含量 GA3 content | ABA含量 ABA content | IAA含量 IAA content | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5.0 cm | 10.0 cm | 5.0 cm | 10.0 cm | 5.0 cm | 10.0 cm | 5.0 cm | 10.0 cm | 5.0 cm | 10.0 cm | 5.0 cm | 10.0 cm | 5.0 cm | 10.0 cm | ||||||||
| 胚芽鞘长度 Coleoptile length | 5.0 | 1.00 | 0.61 | 0.32 | -0.54 | 0.69 | -0.24 | 0.59 | |||||||||||||
| 10.0 | 1.00 | 0.93 | 0.37 | 0.42 | 0.79 | 0.46 | 0.84 | ||||||||||||||
| α-AMY活性 α-AMY activity | 5.0 | 0.61 | 1.00 | 0.68 | 0.09 | 0.99 | 0.60 | 0.99 | |||||||||||||
| 10.0 | 0.93 | 1.00 | 0.57 | 0.57 | 0.94 | 0.28 | 0.97 | ||||||||||||||
| ADH活性 ADH activity | 5.0 | 0.32 | 0.68 | 1.00 | 0.62 | 0.71 | 0.66 | 0.68 | |||||||||||||
| 10.0 | 0.37 | 0.57 | 1.00 | 0.99 | 0.80 | 0.51 | 0.74 | ||||||||||||||
| ROS含量 ROS content | 5.0 | -0.54 | 0.09 | 0.62 | 1.00 | 0.06 | 0.78 | 0.11 | |||||||||||||
| 10.0 | 0.42 | 0.57 | 0.99 | 1.00 | 0.79 | 0.61 | 0.73 | ||||||||||||||
| GA3含量 GA3 content | 5.0 | 0.69 | 0.99 | 0.71 | 0.06 | 1.00 | 0.53 | 0.99 | |||||||||||||
| 10.0 | 0.79 | 0.94 | 0.80 | 0.79 | 1.00 | 0.32 | 0.99 | ||||||||||||||
| ABA含量 ABA content | 5.0 | -0.24 | 0.60 | 0.66 | 0.78 | 0.52 | 1.00 | 0.62 | |||||||||||||
| 10.0 | 0.46 | 0.28 | 0.51 | 0.61 | 0.32 | 1.00 | 0.31 | ||||||||||||||
| IAA含量 IAA content | 5.0 | 0.59 | 0.99 | 0.68 | 0.11 | 0.99 | 0.62 | 1.00 | |||||||||||||
| 10.0 | 0.84 | 0.97 | 0.74 | 0.73 | 0.99 | 0.31 | 1.00 | ||||||||||||||
3 讨论
水稻直播是一种轻简、高效、节水和省工的栽培方式,但由于田间的不良淹水导致O2浓度过低,使出苗率急剧下降,严重影响直播稻的发展[6-7]。研究[2,25]表明,种子萌发所需的O2浓度低于10%时,种子出苗率会显著降低。因此,选择耐低氧萌发能力强的水稻品种是解决这一问题的关键措施。研究[9-10]表明,在淹水萌发时,水稻品种间胚芽鞘长度存在着明显的差异,可将胚芽鞘长度作为耐淹水萌发的指标。Sun等[26]以10 cm水深处理的胚芽鞘长度为指标进行萌发耐淹性鉴定,在粳型杂草稻中克隆了决定低氧下强萌发的关键基因OsGF14h。王洋等[27]以5 cm水深下的胚芽鞘长度为指标进行耐低氧萌发测定,在2个水稻定位群体中检测到8个耐缺氧能力的QTL。综上,5或10 cm淹水深度可作为筛选水稻耐低氧萌发的处理条件,胚芽鞘长度可作为耐淹水萌发的指标。本研究通过0.5(CK)、5.0和10.0 cm淹水处理,以胚芽鞘长度和种子存活率为耐淹指标,对432份水稻种质资源进行低氧萌发筛选。种子耐低氧萌发在品种间存在着差异[10,16,28]。淹水处理下,432份水稻种质资源的胚芽鞘长度和种子存活率表现出广泛的变异,与前人[28]研究结果一致。本研究中,淹水处理下,胚芽鞘长度与种子存活率有极显著正相关关系。经鉴定和筛选得到10个在低氧条件下萌发状况良好的水稻品种,这些资源可作为亲本材料用来选育耐低氧萌发的水稻品种。综上所述,本研究以5.0和10.0 cm淹水处理下胚芽鞘长度作为耐淹水萌发的指标所筛选出的耐淹水种质材料是可靠的。
淹水胁迫下,GA3、α-AMY和IAA协同促进胚芽鞘细胞伸长,使胚芽鞘快速生长,从而帮助水稻幼苗尽快突破水面。在水稻种子萌发过程中,α-AMY是催化淀粉水解的关键酶。当水稻种子处于淹水缺氧环境时,α-AMY活性增强,加速分解胚乳中的淀粉,为种子萌发和幼苗生长提供能量和碳源,保证水稻在淹水条件下有足够的营养物质来支持其生命活动,从而提高水稻的耐淹萌发能力[21]。IAA对水稻种子的萌发和幼苗生长具有重要的调节作用。在淹水条件下,IAA通过调节细胞壁松弛相关基因的表达,使细胞壁松弛,细胞更容易吸水膨胀,进而促进细胞伸长,最终导致胚芽鞘伸长[19]。GA3能打破水稻种子的休眠,促进胚的生长和发育,加速种子萌发。在淹水条件下,GA3可通过提高α-AMY等水解酶的活性,同时通过降解DELLA蛋白促进IAA合成,从而提高水稻种子在淹水逆境下的萌发率[19-20]。本研究中,耐低氧萌发品种的α-AMY活性、GA3和IAA含量在5.0和10.0 cm淹水处理下都有较好的相关性(r>0.9),且5.0和10.0 cm淹水处理下α-AMY活性、GA3和IAA含量均显著提高。综上所述,GA3作为打破种子缺氧休眠的启动信号,同时协同调节α-AMY活性和IAA含量,与α-AMY活性和IAA含量共同在淹水处理下水稻的萌发中发挥更加重要的作用。
研究[12,29-
4 结论
从432份种质资源中筛选获得10份耐低氧萌发能力强的种质资源,其中粳稻品种葡萄黄和黄皮糯、籼稻品种饿死牛和博B表现出更强的耐低氧萌发能力。相对于ABA、ROS含量和ADH活性,α-AMY活性、GA3和IAA含量在淹水处理下水稻的耐低氧萌发中发挥更加重要的作用,其也可能是影响粳稻和籼稻耐低氧萌发能力差异的主要因素。
参考文献
Performance of dry direct-seeded rice in response to genotype and seeding rate
Current status and challenges of rice production in China
An integrated approach to weed management practices in direct-seeded rice under zero- tilled rice-wheat cropping system
Lower global warming potential and higher yield of wet direct-seeded rice in central China
Physiological mechanisms of flooding tolerance in rice: transient complete submergence and prolonged standing water
The molecular regulatory pathways and metabolic adaptationin the seed germination and early seedling growth of rice in response to low O2 stress
Herbicide programs for control of glyphosate resistant volunteer corn in glufosinate-resistant soybean
iTRAQ-based proteomics investigationof critical response proteins in embryo and coleoptile duringrice anaerobic germination
QTLs associated with tolerance of flooding during germination in rice (Oryza sativa L.)
UDP-glucosyltransferase OsUGT75A promotes submergence tolerance during rice seed germination
Rice seed germination priming by salicylic acid and the emerging role of phytohormones in anaerobic germination
Abscisic acid promotes jasmonic acid biosynthesis via a 'SAPK10-bZIP72-AOC' pathway to synergistically inhibit seed germination in rice (Oryza sativa)
Active oxygen species and antioxidants in seed biology
Seed birth to death: Dual functions of reactive oxygen species in seed physiology
Expression of a gene encoding mitochondrial aldehyde dehydrogenase in rice increases under submerged conditions
Regain flood adaptation in rice through a 14-3- 3 protein OsGF14h
QTL mapping and confirmation for tolerance of anaerobic conditions during germination derived from the rice landrace Ma-Zhan Red
太湖流域水稻种子活力和耐缺氧能力遗传变异研究
Flooding stress: acclimations and genetic diversity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}