
作物杂志,2026, 第3期: 216–224 doi: 10.16035/j.issn.1001-7283.2026.03.028
淹水深度对水稻胚芽鞘伸长的影响及其生理机制
刘冬1( ), 李君霞1, 马梦娟1, 张骐飞2, 李明1, 付景1, 王越涛1, 王付华1, 尹海庆1, 王亚1()
), 李君霞1, 马梦娟1, 张骐飞2, 李明1, 付景1, 王越涛1, 王付华1, 尹海庆1, 王亚1()
- 1
河南省农业科学院粮食作物研究所, 450002, 河南郑州
2方城县气象局, 473200, 河南南阳
Effects of Flooding Depth on Coleoptile Growth in Rice and the Underlying Physiological Mechanisms
Liu Dong1(), Li Junxia1, Ma Mengjuan1, Zhang Qifei2, Li Ming1, Fu Jing1, Wang Yuetao1, Wang Fuhua1, Yin Haiqing1, Wang Ya1()
- 1
Cereal Crops Research Institute ,Henan Academy of Agricultural Sciences Zhengzhou 450002, Henan, China
2Fangcheng Meteorological Service Nanyang 473200, Henan, China
摘要:
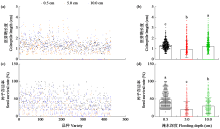
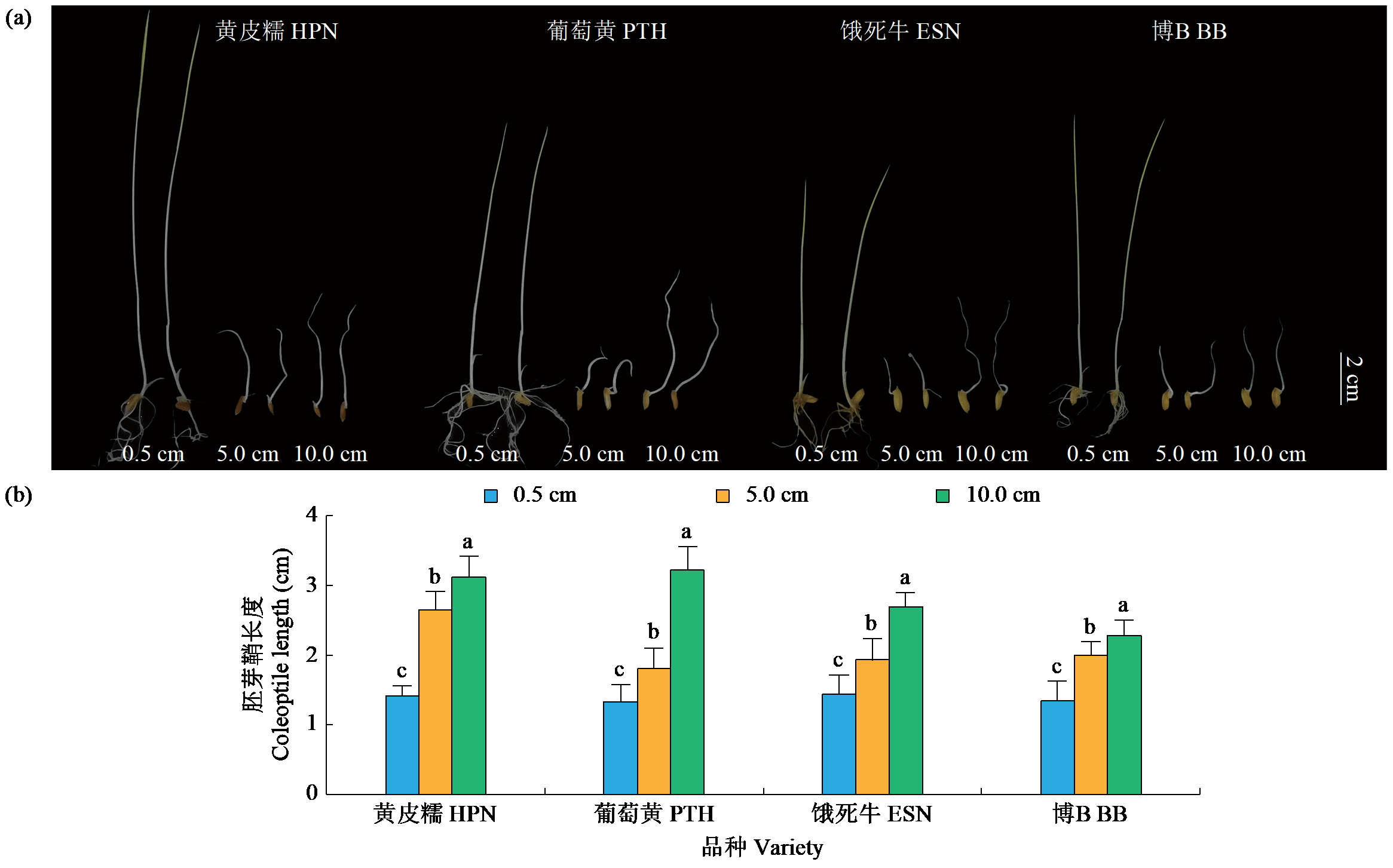
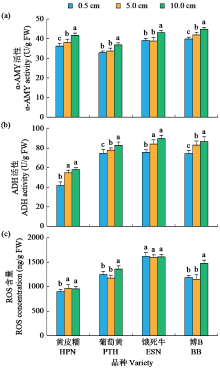
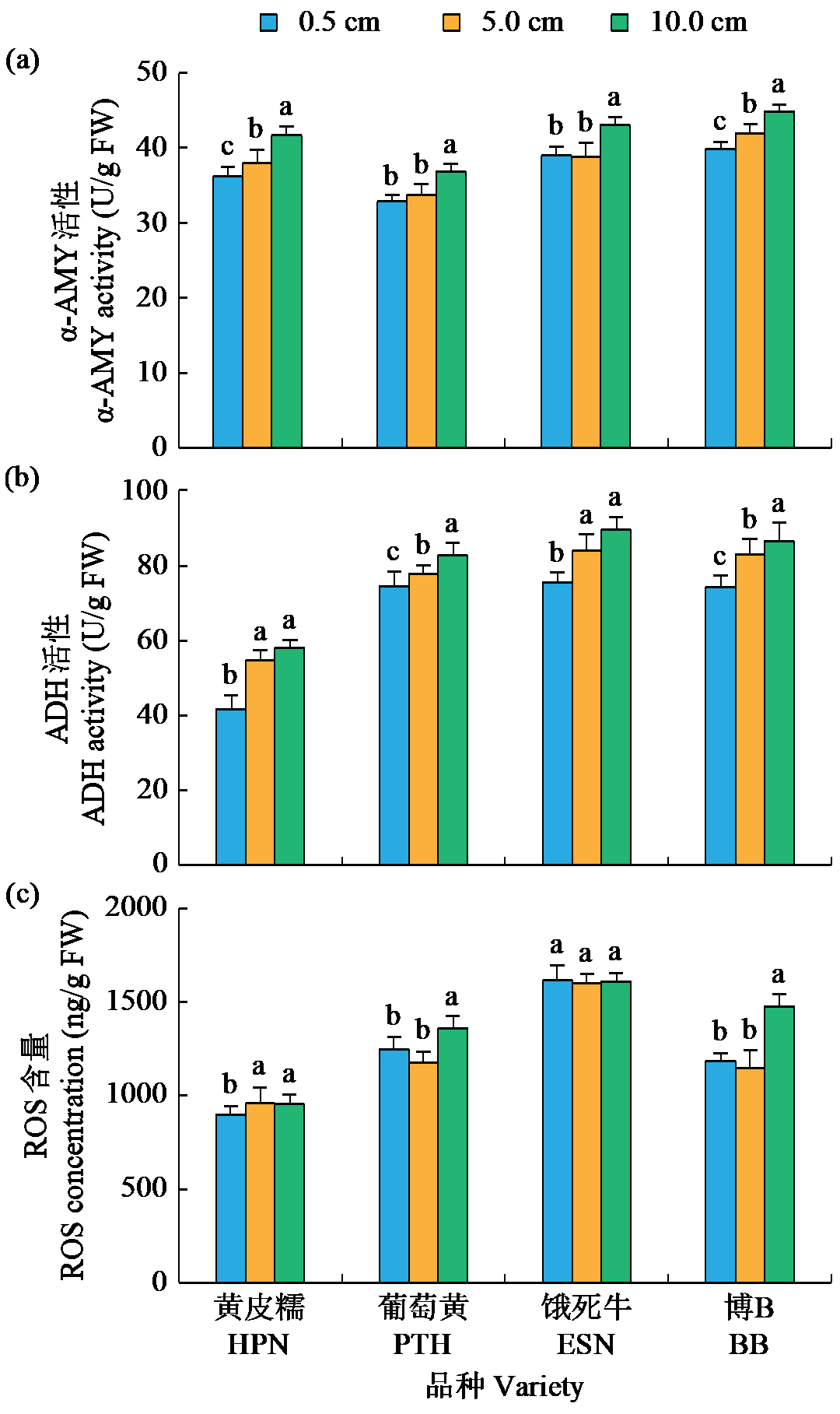
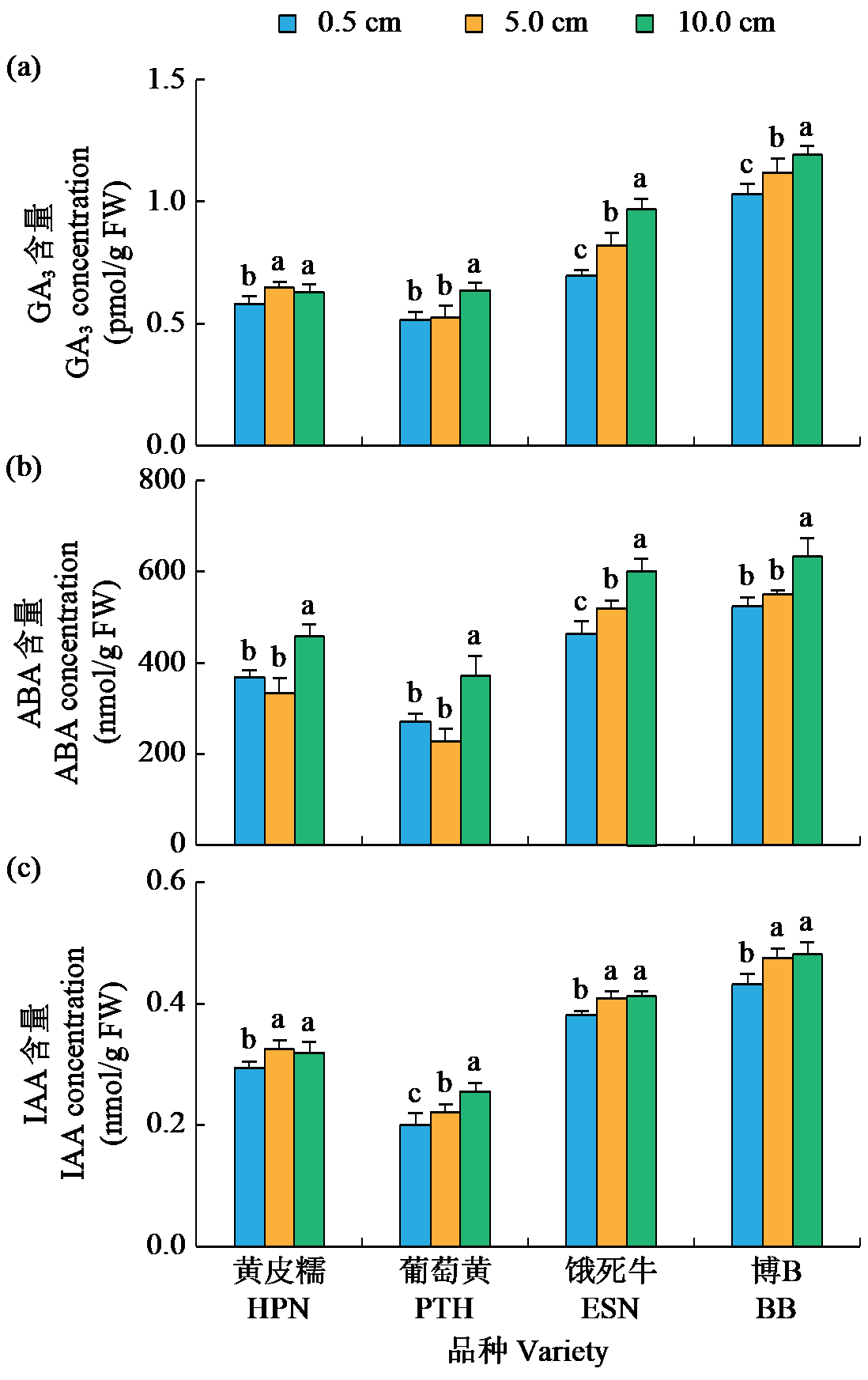
耐低氧萌发种质资源的筛选、利用和生理机制的探究是选育适宜直播水稻新品种的基础。为简便、高效地评价种质资源的耐低氧萌发能力,在0.5(对照)、5.0和10.0 cm淹水深度下,以胚芽鞘长度为主要筛选指标对432份水稻种质资源进行萌发耐淹性鉴定。结果表明,淹水处理下,胚芽鞘长度和种子存活率表现出广泛的变异,且粳稻品种表现出比籼稻品种更长的胚芽鞘。通过胚芽鞘长度和耐淹指数筛选获得10份耐低氧萌发能力强的种质资源。其中,粳稻品种葡萄黄和黄皮糯以及籼稻品种饿死牛和博B表现出更强的耐低氧萌发能力,淹水处理下粳稻品种葡萄黄和黄皮糯的胚芽鞘长度显著高于籼稻品种饿死牛和博B。生理指标表明,淹水处理下4个品种的脱落酸(ABA)、赤霉素3(GA3)和生长素(IAA)含量以及α-淀粉酶(α-AMY)和乙醇脱氢酶(ADH)的活性均显著高于对照处理(0.5 cm),且粳稻品种的α-AMY活性、GA3和IAA含量高于籼稻品种。相关性分析表明,淹水处理下4个品种的α-AMY活性、GA3和IAA含量与胚芽鞘长度呈正相关(r>0.9)。综上,ABA、GA3、IAA和活性氧含量以及α-AMY和ADH活性会促进淹水下胚芽鞘的伸长,而α-AMY活性、GA3和IAA含量在淹水处理下胚芽鞘的伸长中发挥更加重要的作用,且更高的α-AMY活性、GA3和IAA含量赋予粳稻品种更强的耐低氧萌发能力。
| [1] | 刘贵富, 陈明江, 李明, 等. 水稻育种行业创新进展. 植物遗传资源学报, 2018, 19(3):416-429. |
| [2] | Mahajan G, Chauhan B S. Performance of dry direct-seeded rice in response to genotype and seeding rate. Agronomy Journal, 2016, 108:257-265. |
| [3] | 张平, 姜一梅, 曹鹏辉, 等. 通过分子标记辅助选择将耐储藏主效QTL qSS-9Kas转入宁粳4号提高其种子贮藏能力. 作物学报, 2019, 45(3):335-343. |
| [4] | Peng S B, Tang Q Y, Zou Y B. Current status and challenges of rice production in China. Plant Production Science, 2009, 12:3-8. |
| [5] | Ghosh D, Singh U P, Brahmachari K, et al. An integrated approach to weed management practices in direct-seeded rice under zero- tilled rice-wheat cropping system. International Journal of Pest Management, 2017, 63:37-46. |
| [6] | Tao Y, Chen Q, Peng S B, et al. Lower global warming potential and higher yield of wet direct-seeded rice in central China. Agronomy for Sustainable Development, 2016, 36(2):24-32. |
| [7] | 王慧. 水稻直播技术特点及发展现状. 农业科技与装备, 2017(1):72-73. |
| [8] | 姜心禄, 杨永波, 付明全, 等. 不同直播方式下种子处理方式对直播稻出苗和产量的影响. 中国稻米, 2017, 23(4):111-114. |
| [9] | 孙凯, 李冬秀, 杨靖, 等. 水稻耐淹成苗率相关性状全基因组的关联分析. 中国农业科学, 2019, 52(3):385-398. |
| [10] | 刘利成, 李小湘, 黎用朝, 等. 水稻种子耐厌氧萌发全基因组关联分析. 植物遗传资源学报, 2021, 22(6):1644-1650. |
| [11] | Colmer T D, Armstrong W, Greenway H, et al. Physiological mechanisms of flooding tolerance in rice: transient complete submergence and prolonged standing water. Progress in Botany, 2014, 75:255-307. |
| [12] | Ma M Q, Cen W J, Li R B, et al. The molecular regulatory pathways and metabolic adaptationin the seed germination and early seedling growth of rice in response to low O2 stress. Plants, 2020, 9(10):1363-1376. |
| [13] | Chahal P S, Jhala A. Herbicide programs for control of glyphosate resistant volunteer corn in glufosinate-resistant soybean. Weed Technology, 2015, 29(3):431-443. |
| [14] | Zhang G C, Liu Z M, Liu Y H, et al. iTRAQ-based proteomics investigationof critical response proteins in embryo and coleoptile duringrice anaerobic germination. Rice Science, 2021, 28(4):391-401. |
| [15] | Angaji S A, Septiningsih E M, Mackill D J, et al. QTLs associated with tolerance of flooding during germination in rice (Oryza sativa L.). Euphytica, 2010, 172:159-168. |
| [16] | 陈振挺, 冯芳君, 严明, 等. 水稻自然变异群体淹水发芽相关特性鉴定. 上海农业学报, 2020, 36(5):1-6. |
| [17] | 孙志广, 王宝祥, 周振玲, 等. 水稻萌发耐淹性种质资源筛选及QTL定位. 作物学报, 2021, 47(1):61-70. |
| [18] | He Y Q, Sun S, Zhao J, et al. UDP-glucosyltransferase OsUGT75A promotes submergence tolerance during rice seed germination. Nature Communications, 2023, 14(1):2296. |
| [19] | He Y Q, Zhao J, Wang Z F. Rice seed germination priming by salicylic acid and the emerging role of phytohormones in anaerobic germination. Journal of Integrative Plant Biology, 2024, 66(8):1537-1539. |
| [20] | Wang Y F, Hou Y X, Qiu J H, et al. Abscisic acid promotes jasmonic acid biosynthesis via a 'SAPK10-bZIP72-AOC' pathway to synergistically inhibit seed germination in rice (Oryza sativa). New Phytologist, 2020, 228(4):1336-1353. |
| [21] | 钱海丰, 赵晓娟, 赵心爱. α-淀粉酶基因表达的调控. 西北农业学报, 2003, 12(4):87-90,123. |
| [22] | Bailly C. Active oxygen species and antioxidants in seed biology. Seed Science Research, 2004, 14(2):93-107. |
| [23] | Kumar S P J, Rajendra P S, Banerjee R, et al. Seed birth to death: Dual functions of reactive oxygen species in seed physiology. Annals of Botany, 2015, 116(4):663-668. |
| [24] | Nakazono M, Tsuji H, Li Y, et al. Expression of a gene encoding mitochondrial aldehyde dehydrogenase in rice increases under submerged conditions. Plant Physiology, 2000, 124:587-598. |
| [25] | 朱鹤健. 土壤地理学. 北京: 高等教育出版社, 1992. |
| [26] | Sun J, Zhang G C, Cui Z B, et al. Regain flood adaptation in rice through a 14-3- 3 protein OsGF14h. Nature Communications, 2022, 13(1):5664. |
| [27] | 王洋, 郭媛, 洪德林. 水稻幼苗耐缺氧能力的QTL分析. 中国水稻科学, 2010, 24(1):18-24. |
| [28] | Septiningsih E M, Ignacio J I, Sendon P D, et al. QTL mapping and confirmation for tolerance of anaerobic conditions during germination derived from the rice landrace Ma-Zhan Red. Theoretical and Applied Genetics, 2013, 126:1357-1366. |
| [29] | 侯名语, 江玲, 王春明, 等. 水稻种子低氧发芽力的QTL定位和上位性分析. 中国水稻科学, 2004, 18(6):483-488. |
| [30] | 王洋, 王盈盈, 洪德林. 太湖流域水稻种子活力和耐缺氧能力遗传变异研究. 南京农业大学学报, 2009, 32(3):1-7. |
| [31] | Bailey-Serres J, Voesenek L A C J. Flooding stress: acclimations and genetic diversity. Annual Review of Plant Biology, 2008, 59:313-339. |
| [1] | 郭威, 孙冬, 闫治霖, 丁蓓, 杜跃辰, 吴慧扬, 徐强, 窦志, 高辉. 灌浆结实期半深水灌溉对水稻产量、干物质转运及籽粒灌浆的影响[J]. 作物杂志, 2026, (3): 21–29 |
| [2] | 张清芮, 凌凤楼, 高子婷, 王玉, 苏庆旺, 韩笑. 不同叶施时期和锌肥种类对水稻产量和品质的影响[J]. 作物杂志, 2026, (3): 38–47 |
| [3] | 王金玲, 潘越, 李思宇, 沈炘垭, 刘立军. 枯草芽孢杆菌和哈茨木霉菌对水稻稻米品质的影响[J]. 作物杂志, 2026, (2): 202–208 |
| [4] | 陆楚盛, 赖嘉俊, 唐偲, 梁开明, 覃元钰, 钟旭华, 潘俊峰, 刘彦卓, 胡香玉, 胡锐, 李妹娟, 王昕钰, 尹媛红, 叶群欢, 沈宏, 傅友强. 铵、硝营养条件下不同酸碱度对水稻苗期生长和养分吸收的影响[J]. 作物杂志, 2026, (2): 90–97 |
| [5] | 孙茹梦, 张男, 殷佳, 汝艳, 景文疆, 张耗. 水稻根系分泌物对干旱胁迫的响应研究进展[J]. 作物杂志, 2026, (1): 1–8 |
| [6] | 刘晴, 孙露宏, 高世伟, 刘宇强, 常汇琳, 马成, 王婧泽, 王翠玲, 聂守军. 水稻叶片的生理性状和形态特征受铬胁迫的影响研究[J]. 作物杂志, 2026, (1): 143–151 |
| [7] | 吴叶宁, 胡昊驰, 王春勇, 谢锐, 张永虎, 温蕊, 金晓蕾. 基于SSR标记的104份苦荞种质资源遗传多样性及群体结构分析[J]. 作物杂志, 2026, (1): 26–32 |
| [8] | 于秋竹, 邱俊华, 杜含梅, 王安虎. 不同苦荞种质资源生长发育及硒响应特征综合评价研究[J]. 作物杂志, 2026, (1): 72–84 |
| [9] | 李文俊, 郭延平, 杨生华, 邵扬. 513份蚕豆种质资源主要农艺性状遗传多样性分析[J]. 作物杂志, 2026, (1): 85–93 |
| [10] | 闸雯俊, 李行润, 周发松, 冯芳, 吴边, 陈俊孝, 石少阶, 周雷, 王静, 游艾青. 水稻光温敏不育系19XS的选育及其特性分析[J]. 作物杂志, 2026, (1): 9–14 |
| [11] | 陈雷, 唐茂艳, 张战营, 钟晓媛, 高国庆, 张晓丽, 梁天锋, 潘英华. 花期高温胁迫下水稻种质资源的外观品质分析与评价[J]. 作物杂志, 2025, (6): 132–139 |
| [12] | 王舒琦, 李建波, 刘志萍, 马宇, 渠佳慧, 巴图, 徐寿军. 不同栽培模式下大麦产量与蛋白质形成的生理机制研究[J]. 作物杂志, 2025, (6): 172–180 |
| [13] | 陈浩楠, 赵宏岩, 张文静, 张琦, 杜吉到, 王琦瑞, 任若然, 韩毅强. 豌豆SSR分子标记筛选与指纹图谱的构建[J]. 作物杂志, 2025, (6): 19–27 |
| [14] | 孙强, 阮辛森, 周志豪, 孙会娟, 徐冉, 凌冬, 赵翠荣. 不同类型水稻品种表型性状遗传多样性分析[J]. 作物杂志, 2025, (6): 28–36 |
| [15] | 杨硕, 马晓娟, 郑璇, 侯艺, 叶林, 程国新. 100份辣椒种质资源遗传多样性分析及抗病性评价[J]. 作物杂志, 2025, (6): 73–82 |
|
||