
作物杂志,2016, 第4期: 75–79 doi: 10.16035/j.issn.1001-7283.2016.04.012
非生物逆境胁迫下普通小麦烟农19幼苗FeSOD基因表达分析
李钰1,郑文寅2,冯春3,王荣富4,李娟4
- 1 安徽农业大学资源与环境学院,230036,安徽合肥
2 安徽农业大学农学院,230036,安徽合肥
3 安徽农业大学农业园,230036,安徽合肥
4 安徽农业大学生物技术中心,230036,安徽合肥
Analysis of FeSOD Gene Expression in Normal Wheat Yannong 19 Seedlings under Abiotic Stress
Li Yu1,Zheng Wenyin2,Feng Chun3,Wang Rongfu4,Li Juan4
- 1 School of Resources and Environment,Anhui Agricultural University,Hefei 230036,Anhui,China
2 C ollege of Agronomy,Anhui Agricultural University,Hefei 230036,Anhui,China
3 Agriculture Demonstration Park Management Center,Anhui Agricultural University,Hefei 230036,Anhui,China
4 Biotechnology Center,Anhui Agricultural University,Hefei 230036,Anhui,China
摘要:
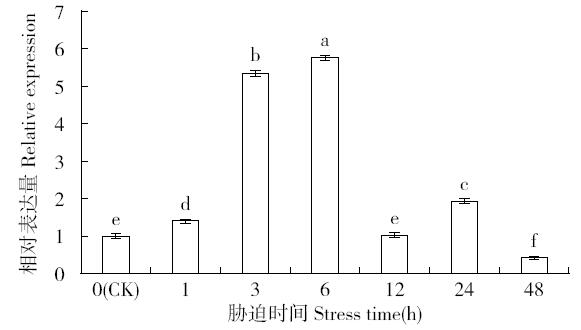
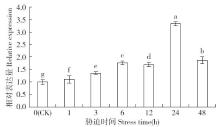
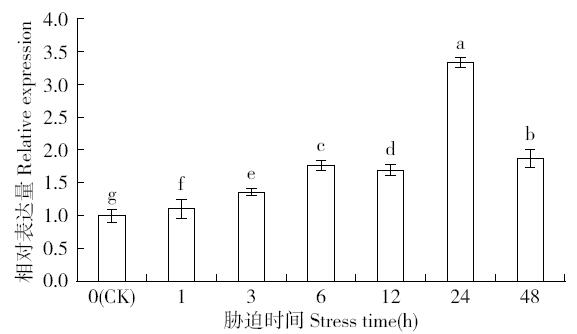
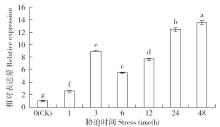
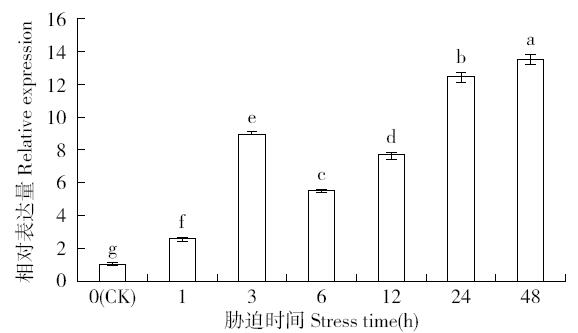
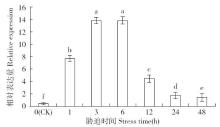
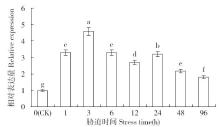
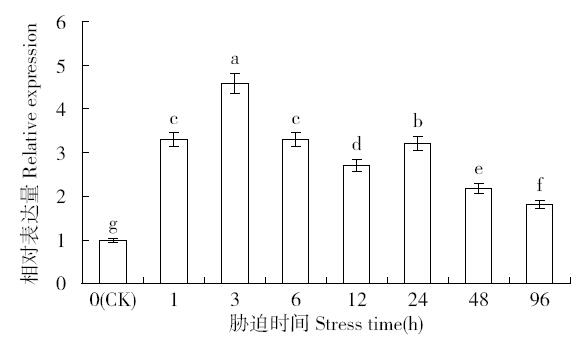
非生物逆境(高温、低温、干旱、盐胁迫等)严重影响作物生长,研究参与逆境胁迫应答基因具有重要的理论意义和应用价值。超氧化物歧化酶(superoxide dismutase,SOD)是一种广泛存在于真核生物中的金属酶类,在植物的抗逆性中起到重要作用。采用qRT-PCR技术,分析普通小麦烟农19幼苗叶中FeSOD基因在盐、脱落酸(ABA)、干旱、高温、低温胁迫过程中的的表达模式。结果表明:在逆境胁迫下,普通小麦烟农19幼苗FeSOD基因的表达量大体呈现先上升后下降的趋势。在37℃高温、-4℃低温、300mmol/L NaCl、30%的PEG-6000和100μmol/L ABA胁迫下,FeSOD基因的表达量分别在3、3、6、48和24h时最高,分别为对照的34.0、4.6、4.3、5.8、13.5和3.3倍,差异均达到显著水平,说明FeSOD基因在普通小麦烟农19幼苗逆境胁迫中发挥着重要的调控功能,为进一步了解小麦抗逆分子机制和改良小麦品种提供理论依据。


| [1] |
Ahuja I Vos R C H D,Bones A M, et al. Plant molecular stress responses face climate change. Trends in Plant Science, 2010,15(12):664-674.
doi: 10.1016/j.tplants.2010.08.002 |
| [2] | Kim K Y, Sang Y L, Cho Y S , et al. Molecular characterization and mRNA expression during metal exposure and thermal stress of copper/zinc- and manganese-superoxide dismutases in disk abalone,Haliotis discus discus. Fish & Shellfish Immunology, 2007,23(5):1043-1059. |
| [3] |
Yu Q, Osborne L D, Rengel Z . Increased tolerance to Mn deficiency in transgenic tobacco overproducing superoxide dismutase. Annals of Botany, 1999,84(4):543-547.
doi: 10.1006/anbo.1999.0951 |
| [4] |
Yu Q, Rengel Z . Drought and salinity differentially influence activities of superoxide dismutases in narrow-leafed lupins. Plant Science, 1999,142(1):1-11.
doi: 10.1016/S0168-9452(98)00246-5 |
| [5] | 田春美, 钟秋平 . 超氧化物歧化酶的现状研究进展. 中国热带医学, 2005,5(8):1730-1732. |
| [6] |
Prabhakar R, Morokuma K, Musaev D G . A DFT study of the mechanism of Ni superoxide dismutase (NiSOD): Role of the active site cysteine-6 residue in the oxidative half-reaction. Journal of Computational Chemistry, 2006,27(12):1438-1445.
doi: 10.1002/jcc.v27:12 |
| [7] |
杨鸯鸯, 李云, 丁勇 , 等. 甘蓝型油菜Cu/ZnSOD和FeSOD基因的克隆及菌核病菌诱导表达. 作物学报, 2009,35(1):71-78.
doi: 10.3724/SP.J.1006.2009.00071 |
| [8] |
Canini A, Albertano P, Caiola M G . Localization of Fe‐containing superoxide dismutase in cyanobacteria from the Baltic Sea: depth and light dependency. New Phytologist, 1998,139(139):247-254.
doi: 10.1046/j.1469-8137.1998.00187.x |
| [9] | 李琳玲, 程华, 许锋 , 等.银杏铁型超氧化物歧化酶基因( GbFeSOD)的克隆与表达.果树学报,2009(6):840-846. |
| [10] |
Ben R I, Lenne C, Leblanc N , et al. Iron-superoxide dismutase and monodehydroascorbate reductase transcripts accumulate in response to internode rubbing in tomato. Comptes Rendus Biologies, 2004,327(7):679-686.
doi: 10.1016/j.crvi.2004.06.002 |
| [11] |
Van C W, Capiau K, Van M M , et al. Enhancement of oxidative stress tolerance in transgenic tobacco plants overproducing Fe-superoxide dismutase in chloroplasts. Plant Physiology, 1996,112(4):1703-1714.
doi: 10.1104/pp.112.4.1703 |
| [12] | 刘录祥, 赵林姝, 梁欣欣 , 等. 基因枪法获得逆境诱导转录因子DREB1A转基因小麦的研究. 中国生物工程杂志, 2003,23(11):53-56. |
| [13] | Van B F, Slooten L, Stassart J M , et al. Overproduction of Arabidopsis thaliana FeSOD confers oxidative stress tolerance to transgenic maize. Plant & Cell Physiology, 1999,40(5):515-523. |
| [14] |
Alscher R G, Erturk N . Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. Journal of Experimental Botany, 2002,53(372):1331-1341.
doi: 10.1093/jexbot/53.372.1331 |
| [15] | 李斌, 郭世荣, 孙锦 , 等. 外源Spd对盐胁迫下黄瓜SAMs基因表达的影响及SAMs蛋白的生物信息学分析. 园艺学报, 2011,38(12):2289-2296. |
| [16] |
Wu G, Wilen R W, Robertson A J , et al. Isolation,chromosomal localization,and differential expression of mitochondrial manganese superoxide dismutase and chloroplastic copper/zinc superoxide dismutase genes in wheat. Plant Physiology, 1999,120(2):513-520.
doi: 10.1104/pp.120.2.513 |
| [17] | 曲妍妍 .番茄叶绿体LesAPX和Cu/Zn-SOD的克隆及功能分析. 泰安:山东农业大学, 2008. |
| [18] |
Neill S J, Desikan R, Clarke A , et al. Hydrogen peroxide and nitric oxide as signalling molecules in plants. Journal of Experimental Botany, 2002,53(372):1237-1247.
doi: 10.1093/jexbot/53.372.1237 |
| [19] | 覃鹏, 刘飞虎, 梁雪妮 .超氧化物歧化酶与植物抗逆性.黑龙江农业科学,2002(1):31-34. |
| [20] | 马旭俊, 朱大海 . 植物超氧化物歧化酶(SOD)的研究进展. 遗传, 2003,25(2):225-231. |
| [21] | 李娟, 杨立新, 郑文寅 , 等. 小麦FeSOD基因的克隆与序列分析. 核农学报, 2013,27(8):1111-1117. |
| [22] | 李娟, 王泽文, 杨利新 , 等. 小麦Fe超氧歧化酶基因的原核表达分析. 西北植物学报, 2013,33(11):2147-2152. |
| [23] |
何敬和, 姚丽 . 小麦Mn-SOD基因的克隆及其在盐胁迫下的表达分析. 麦类作物学报, 2010,30(4):630-633.
doi: 10.7606/j.issn.1009-1041.2010.04.008 |
| [24] | 李合生 . 现代植物生理学.北京: 高等教育出版社, 2012. |
| [25] | Kwon S I, An C S . Cloning and expression of mitochondrial MnSOD from the small radish (Raphanus sativus L.). Molecules & Cells, 2003,16(2):194-200. |
| [26] | 郁万文, 曹福亮, 汪贵斌 , 等. 银杏抗氧化酶系统对短期梯度变温的响应. 江西农业大学学报, 2008,30(2):252-256. |
| [1] | 刘建霞,张晓丹,王润梅,周凤,刘文英,刘支平. 6-BA浸种对盐胁迫下绿豆萌发及幼苗生理特性的影响[J]. 作物杂志, 2018, (1): 166–172 |
| [2] | 刘畅,李雪妹,谭佳缘,梁晓敏,李雪梅. 聚乙二醇(PEG)模拟水分胁迫对水稻幼苗矿质离子含量的影响[J]. 作物杂志, 2017, (5): 162–167 |
| [3] | 高宇,崔世茂,宋阳,孙世君. CO2加富对温室辣椒幼苗生长及光合特性的影响[J]. 作物杂志, 2017, (5): 80–84 |
| [4] | 王文浩,郑洪元,刘文俊,何丽芬,闫玉星. 外源NO对向日葵种子萌发和幼苗生长的影响[J]. 作物杂志, 2017, (4): 169–172 |
| [5] | 武俊英,秦丽,杨进,秦德志. 盐胁迫对农大甜研6号甜菜幼苗生长和养分运移的研究[J]. 作物杂志, 2017, (3): 75–80 |
| [6] | 张紫薇,庞春花,张永清,倪瑞军,杨世芳,王璐瑗,刘丽琴. 等渗NaCl和PEG胁迫及复水处理对藜麦种子萌发及幼苗生长的影响[J]. 作物杂志, 2017, (1): 119–126 |
| [7] | 孙阳,王燚,孟瑶,樊海潮,曲丹阳,李晶,魏湜,顾万荣. 外源5-氨基乙酰丙酸对低温胁迫下玉米幼苗生长及光合特性的影响[J]. 作物杂志, 2016, (5): 87–93 |
| [8] | 桑利敏,李彩凤,王玉波,刘磊,郭广昊,郭剑,陈明. Na2SO4+NaCl混合盐胁迫对甜菜幼苗生长的影响[J]. 作物杂志, 2016, (4): 137–141 |
| [9] | 卢峰, 张飞. 多水胁迫对高粱幼苗的生理影响[J]. 作物杂志, 2015, (4): 92–96 |
| [10] | 王泳超, 顾万荣, 魏湜, 等. 新型植物生长复配剂对玉米幼苗形态建成及叶片生理特性的调控效应[J]. 作物杂志, 2015, (2): 89–94 |
| [11] | 田鑫, 钟程, 李性苑, 等. 盐胁迫对薏苡种子萌发及幼苗生长的影响[J]. 作物杂志, 2015, (2): 140–143 |
| [12] | 吴文荣, 牛瑞明, 苑莹, 等. 外源NO对模拟干旱胁迫下亚麻种子发芽及幼苗生长的影响[J]. 作物杂志, 2015, (1): 143–147 |
| [13] | 赵俊香, 刘守伟, 吴凤芝. 盐碱胁迫对4种菊芋材料种子萌发及幼苗生长的影响[J]. 作物杂志, 2015, (1): 133–137 |
| [14] | 朱双艳, 齐军仓, 廖乐, 等. 水分胁迫对大麦幼苗叶片表皮蜡质沉积的影响[J]. 作物杂志, 2014, (5): 64–67 |
| [15] | 孙惠娟, 王超. 结球甘蓝幼苗期耐寒性鉴定[J]. 作物杂志, 2014, (3): 58–63 |
|