


水稻是我国最主要的粮食作物之一。依据植物学分类,我国种植的水稻品种属于亚洲栽培稻(Oryza sativa L.),包括粳稻(O. sativa L. ssp. japonica)和籼稻(O. sativa L. ssp. indica)2个亚种[1],它们在形态、发育与生理等方面都表现出较大的差异。粳稻产量往往不及籼稻,但大都比籼稻具有更强的低温耐受性。尽管全球气候总体变暖,但近年来极端气候频繁,如在我国南方籼稻种植地区倒春寒和寒露风等低温灾害常有发生,导致水稻产量损失严重。将粳稻耐低温基因导入籼稻,是改良籼稻抗寒性的有效途径。
借助现代分子生物学技术,人们对于粳稻具有的低温耐受性有了一定的认识,但耐低温胁迫是一个复杂性状,其遗传机理研究进展相对缓慢。据统计,在水稻12条染色体上,超过250个耐低温QTL被定位[2,3,4,5,6,7,8,9,10],然而,大都停留在初定位阶段,精细定位和克隆的基因较少。目前已完成精细定位的QTL约12个,涉及萌发期耐低温的1个(qLTG-9)、苗期耐低温的5个(qCTS4、qCtss11、qSCT1、qSCT11、qLOP2/qPSR2-1)、孕穗期耐低温的5个(qCTB8、qCTB7、qCT-3-2、qLTB3、qCTB10-2)、苗期和成熟期耐低温的1个(qRC10-2)[11]。克隆并进行功能研究的耐低温基因仅有9个,包括Ctb1[12]、GSTZ2[13]、qLTG3-1[14]、LTG1[15]、COLD1[16]、qCTS-9[17]、CTB4a[18]、bZIP73[19]和HAN1[20]等。其中,COLD1基因编码一个G蛋白信号调节因子,其与G蛋白α亚基RGA1互作以感知低温,激活Ca2+通道,以增强G蛋白的鸟苷三磷酸酶(guanosine triphosphatase,GTP)活性,从而提高水稻的耐寒性能。粳型COLD1Jap较籼型COLD1Ind具有更强的低温耐受性,有重要的育种利用价值[16]。将粳型COLD1Jap基因导入籼稻品种,可以提高籼稻的耐寒性。基于粳型COLD1Jap与籼型COLD1Ind存在的SNP2位点DNA序列差异,杨佳等[21]开发了用于鉴定该基因的衍生型酶切扩增多态性序列(derived cleaved amplified polymorphic sequence,dCAPS)的功能标记。然而,dCAPS功能标记需使用限制性内切酶,且扩增片段较小,需采用聚丙烯酰胺凝胶电泳才能取得较好的基因型鉴定效果,这使得在实际应用中存在费用昂贵、操作繁琐及费时费工等缺点。本研究根据扩增受阻突变体系PCR技术原理[22],结合COLD1基因在籼、粳稻功能区存在的序列差异,设计检测该基因的新标记,旨在建立快捷、准确及价格低廉的基因分型法来鉴定COLD1基因的籼粳属性,为该基因的分子标记辅助育种及资源鉴定提供高效分子标记。
1 材料与方法
1.1 供试材料
供试籼稻品种:9311、特青、明恢63、珍汕97和南京6号;粳稻品种:日本晴、越光、空育131、稻花香2号和松粳9号;籼粳杂交F1:9311/越光。籼粳杂交F2群体:南京6号/稻花香2号。
1.2 试验方法
1.2.1 功能标记的设计 水稻籼、粳亚种在COLD1基因第4外显子处存在1个功能单碱基差异SNP2。在低温敏感的籼稻品种中,该碱基为T或C,而在耐低温的粳稻品种中该碱基为A。从NCBI中下载相应的DNA序列(登录号:AP014960↘BAS90836,国家水稻数据中心:http://www.ricedata.cn/gene/list/2781.htm),结合扩增受阻突变体系PCR技术原理进行功能标记设计。功能标记由5条引物组成,即正反向外引物各1条,正向内引物2条,反向内引物1条。
1.2.2 DNA的提取 取水稻的新鲜叶片约0.4g,按照传统的CTAB方法进行DNA提取。
1.2.3 DNA的PCR扩增及产物检测 仅检测COLD1基因的籼粳性,即耐低温性与不耐低温性。PCR扩增体系为20μL:DNA 2μL(10~100ng/L),引物Primer混合液2μL(正反向外引物各2μmol/L,两正向内引物各4μmol/L,反向内引物8μmol/L),10×Taq Buffer 2μL,Mg2+ Buffer 1.2μL,dNTP Mixture 0.4μL(2.5mmol/L),Taq DNA聚合酶0.4μL(2.5U/μL),ddH2O 12μL。
当检测到为籼型COLD1Ind时,若需进一步鉴定其差异性位点碱基(T或C),则只需将上述引物混合液组分构成更改为2μL(正反向外引物各2μmol/L,任一正向内引物4μmol/L,反向内引物4μmol/L)。
反应程序:94℃ 5min;94℃ 30s,60℃ 30s,72℃1min,循环31次;在72℃延伸10min,10℃ 1min,产物取出备用。
电泳检测:扩增产物在约2%的琼脂糖凝胶中进行电泳检测。
2 结果与分析
2.1 COLD1基因功能标记的设计
根据基因在籼型COLD1Ind为碱基T或C,在粳型COLD1Jap为碱基A,结合扩增受阻突变体系PCR技术原理,设计内外向引物。外引物用斯坦福大学的在线引物设计软件进行设计,正反引物均位于保守区域编码区,正向外引物用COLD1-O-F表示,反向外引物用COLD1-O-R表示;内引物由人工设计,3′端落在SNP差异位点上,检测粳型COLD1Jap的反向内引物为COLD1-I-R。由于籼型的COLD1Ind在SNP位点上是碱基T或C,因此,检测籼型COLD1Ind的正向内引物有2条,即COLD1-I-F1和COLD1-I-F2;为增强特异性,在3条内引物的3′端的第3个碱基各引入1个错配碱基。上述引物设计策略见图1。
图1
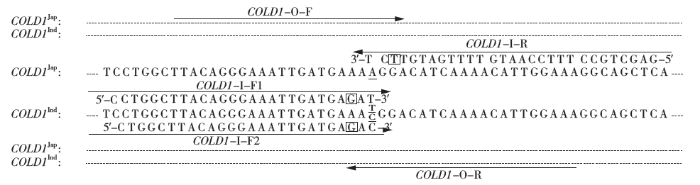
图1
COLD1基因功能标记设计策略
序列中字母下划线表示单核苷酸差异位点SNP2;省略号表示相同的碱基;箭头表示引物位置和扩增方向;方框内为引入的错配碱基
Fig.1
Design strategies for the primers to detect COLD1 gene
The underlines indicate the functional polymorphism SNP2 in the COLD1 gene; the ellipses indicate the same base; the arrows indicate the position and amplification direction of the corresponding primers; the letters in the box present the mismatched bases
根据引物设计策略,5条引物在同一PCR反应体系里进行扩增。理论预计:所有水稻品种均可由外引物COLD1-O-F和COLD1-O-R扩增出长度为965bp的片段,并作为阳性对照;含纯合粳型COLD1Jap水稻品种可由正向外引物COLD1-O-F和反向内引物COLD1-I-R扩增出长度为458bp的片段;含纯合籼型COLD1Ind水稻品种能扩增出长度为562bp(正向内引物COLD1-I-F1和反向外引物COLD1-O-R)或561bp(正向内引物COLD1-I-F2和反向外引物COLD1-O-R)的片段;而籼粳杂合子可扩增出965、562(或561)及458bp 3种类型的条带。引物序列如表1所示。
表1 COLD1基因功能标记的引物序列
Table 1
| 引物Primer | 序列(5′-3′)Sequence (5′-3′) |
|---|---|
| COLD1-O-F | CATTTCCCCATGCCTTCTCC |
| COLD1-O-R | CAACTGTCCCAACGATACGC |
| COLD1-I-F1 | CCTGGCTTACAGGGAAATTGATGAGAT |
| COLD1-I-F2 | CTGGCTTACAGGGAAATTGATGAGAC |
| COLD1-I-R | GAGCTGCCTTTCCAATGTTTTGATGTTCT |
2.2 COLD1基因功能标记验证
选用Ma等[16]对COLD1基因已经测序的9311、特青、明恢63、珍汕97、南京6号、日本晴、越光、空育131、稻花香2号、松粳9号及F1(9311/越光)等11个材料对功能标记进行验证。将所设计的5个引物在同一PCR反应体系里对上述水稻材料进行扩增。电泳图谱(图2)显示,日本晴、越光、空育131、稻花香2号和松粳9号粳稻品种扩增出了约458bp的片段,特异性扩增COLD1基因编码区SNP2位点的核苷酸碱基A,为正向外引物COLD1-O-F和反向内引物COLD1-I-R扩增的产物;9311、特青、明恢63、珍汕97及南京6号5个籼稻品种扩增出了约562bp的片段,特异性扩增COLD1基因编码区SNP2位点的核苷酸碱基T,为正向内引物COLD1-I-F1和反向外引物COLD1-O-R所扩增。籼粳F1扩增出了以上籼粳品种的所有特征性条带(图2)。供试材料均得到了有效扩增,且所扩增条带的类型与测序的碱基差异相吻合。
图2
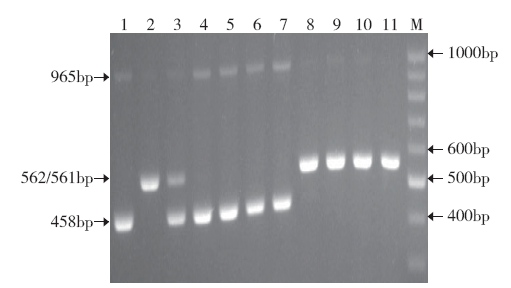
图2
COLD1基因功能标记对水稻材料的检测
M:Ladder H1(100、200、300、400、500、600、700、800、900、1000bp),下同;1-11分别为日本晴、9311、F1(9311/越光)、越光、空育131、稻花香2号、松粳9号、特青、明恢63、珍汕97和南京6号
Fig.2
Detection of rice materials by functional marker of COLD1 gene
M: Ladder H1 (100, 200, 300, 400, 500, 600, 700, 800, 900, 1000bp),the same below; 1-11 are Nipponbare, 9311, F1 (9311/Koshihikari), Koshihikari, Kongyu 131, Daohuaxiang 2, Songjing 9, Teqing, Minghui 63, Zhenshan 97, and Nanjing 6, respectively
COLD1-I-F2引物的作用没有得到体现,那是由于该引物针对检测SNP2位点的差异碱基C,而所检测的水稻材料不存在与之相应的差异碱基的缘故。上述结果表明所设计的功能标记可以对粳型COLD1Jap和籼型COLD1Ind及其杂合子进行鉴定区分。理论上,所有的供试材料还应出现由正向外引物COLD1-O-F和反向外引物COLD1-O-R扩增的大小为965bp的条带,然而,从图2不难看出,有的材料仅出现了微弱的长度为965bp的条带,有些材料甚至没有显示该条带,很可能是由于在多引物扩增的反应体系中,存在明显的竞争效应,小片段扩增所需条件相对较低,获得扩增优势,扩增效率高,而与之相反,大片段处于扩增劣势,扩增效率低,产物量少,条带不明显。值得指出的是,965bp条带为所有材料共有条带,不属于特征条带,其是否出现,并不影响对COLD1基因籼粳属性的鉴定。
2.3 COLD1基因功能标记对南京6号/稻花香2号F2个体的基因型鉴定
利用COLD1基因的引物功能标记对F2(南京6号/稻花香2号)群体进行SNP2位点基因型检测。从F2代群体中随机选择112株水稻植株,用COLD1基因功能标记进行鉴定。检测结果表明,在该F2群体被检测材料中,33株出现COLD1Ind基因型特征条带(562bp),25株出现COLD1Jap基因型特征条带(458bp),54株出现杂合型特征条带(458bp和562bp),部分植株电泳图谱见图3。在F2群体中,籼型COLD1Ind、杂合型COLD1Ind/COLD1Jap、粳型COLD1Jap 3种基因型的分离比例符合1:2:1(χ2=1.286),进一步证实了本研究中所设计的COLD1基因新功能标记能准确鉴定该基因3种不同的类型,可用于鉴定相关的资源和标记辅助选择育种。
图3
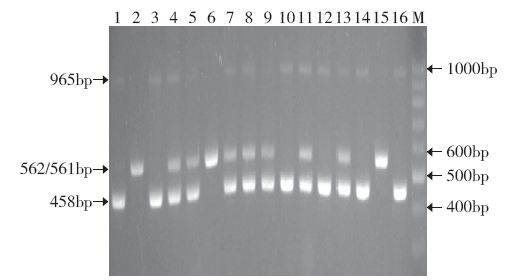
图3
使用功能标记对南京6号/稻花香2号F2群体16个植株的检测
Fig.3
Identification of the genotypes of SNP2 site at COLD1 in 16 F2 individuals from the cross Nanjing 6/Daohuaxiang 2 using functional marker
2.4 籼型COLD1Ind基因SNP2位点突变碱基的鉴定
研究证实,COLD1基因的第4外显子区域SNP2的突变位点在粳型COLD1Jap中为碱基A,而在籼型COLD1Ind中为碱基T或C,导致了籼粳材料对低温耐受性的差异。本研究根据这些碱基的差异,设计了5个引物组成的功能标记对目的基因进行鉴定。在5条引物中,其中3条内引物是针对3种不同差异碱基而设计的,COLD1-I-R是鉴定粳型COLD1Jap碱基A的特异性内引物;而COLD1-I-F1和COLD1-I-F2分别是鉴定籼型COLD1Ind中碱基T和C的特异性引物。一般只需将5条引物混合在同一PCR反应体系中扩增来对COLD1基因的籼、粳型进行鉴定,无论籼型COLD1Ind中的SNP2是碱基T或C,在约560bp(562bp或561bp)处均有条带产生,能很好地与特征带为458bp的粳型COLD1Jap区分。
在5种引物混合鉴定确定是籼型COLD1Ind后,若要具体鉴定差异碱基是T或C,则将鉴定该位点的特征内引物COLD1-I-F1或COLD1-I-F2分别与另外3条引物组成4条引物在同一PCR反应体系扩增即可(图4)。从图4中可看出,在COLD1-I-F1参与的4条引物中,籼型COLD1Ind中SNP2位点为碱基T的9311扩增出了562bp的特征条带,与预测扩增片段大小一致(图4泳道5);而COLD1-I-F2参与的4条引物中,由于9311中SNP2位点不是碱基C,因而没有扩增出561bp的特征条带(图4泳道2),这说明COLD1-I-F1和COLD1-I-F2这两个内引物能独立且准确地鉴定籼型COLD1Ind中SNP2位点具体碱基类型T或C(即在有引物COLD1-I-F1参与的反应体系中,SNP2位点碱基类型为T,可扩增出562bp的条带,而碱基类型是C的无此条带;同理,在COLD1-I-F2参与的反应体系中,SNP2位点碱基类型为C,可扩增出561bp的条带,而碱基类型是T的无此条带)。此外,从图4泳道编号为2中还能看出,由于内引物没有得到有效扩增,竞争效应消除,两条外引物组成的大片段(965bp)能进行较好的扩增,验证了上文提及的竞争效应,即小片段优先获得扩增优势的猜测。
图4
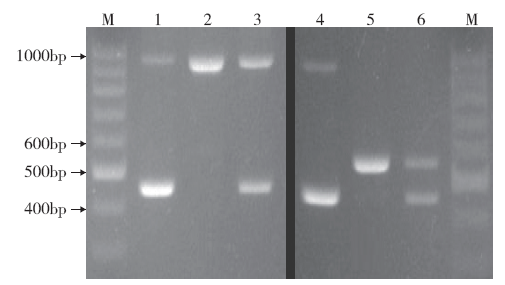
图4
使用4条引物对籼型COLD1Ind基因差异碱基的鉴定
1和4为越光;2和5为9311;3和6为F1(9311/越光)
Fig.4
Identification of differential bases of COLD1Ind gene in Indica rice by four primers
1 and 4: Koshihikari; 2 and 5: 9311; 3 and 6: F1 (9311/Koshihikari)
以上试验结果表明,本研究所开发的新功能标记不需用限制性内切酶,不仅能简便快速地对COLD1的籼粳属性进行鉴定,还弥补了不能对籼型COLD1Ind的SNP2位点具体碱基类型鉴定[21]的不足。
3 讨论
近年来,水稻受到低温冷害导致严重减产的事件常有发生,特别是籼稻对低温更为敏感。鉴于此,人们加大了对低温冷害的关注,如在贵州,耐低温性鉴定成为该省籼稻品种审定的一个重要环节,所有参加区试的籼稻新品系(组合)均需在几个海拔较高的冷凉粳稻区进行耐低温性鉴定。随着分子生物学的不断发展,凭借分子技术手段,对耐低温基因的研究取得了较大的进展,特别是近几年,通过籼粳的低温耐受性对比研究,一些重要的耐低温基因被挖掘,为育种利用提供前提和基础条件。
已有的研究[16]表明,COLD1是一个优秀的耐冷性基因,粳型基因COLD1Jap具有较好的耐冷性能,可将COLD1Jap导入籼稻,提高籼稻耐冷性。功能标记是属于共分离的分子标记,依据基因本身的DNA序列差异而设计,因此,能准确鉴定出目标等位基因,高效率地对目标基因进行鉴定筛选,是分子标记中最为理想的标记类型。杨佳等[21]依据COLD1基因在籼粳间存在单核苷酸差异特性,开发了基于酶切的功能分子标记,能对COLD1基因的籼粳类型进行准确鉴定。然而,在实践应用中,该标记还存在一些不足,一是涉及内切酶的应用费用较高;二是扩增片段较小,不适合操作简便的琼脂糖凝胶电泳检测,而聚丙烯酰胺凝胶操作复杂;三是不能对籼型COLD1基因的具体突变碱基类型进行鉴定等。鉴于COLD1基因在育种中具有重要的应用价值,有必要开发操作简便高效且费用低廉的替代标记,更好地促进该基因在育种中的推广应用。本研究基于扩增受阻突变体系PCR技术原理,结合COLD1基因在籼粳的第4条染色体第4外显子SNP2存在的碱基差异,粳型COLD1Jap为A,而籼型COLD1Ind为T或C,设计了针对3个不同碱基差异的特异性内引物,其中,COLD1-I-R为粳型COLD1Jap特异性内引物,其与正向外引物COLD1-O-F引物组成标记,扩增458bp大小的特征条带;COLD1-I-F1、COLD1-I-F2分别为籼型碱基T和C的特异性内引物,与反向外引物COLD1-O-R组成标记,扩增出562bp或561bp的特征条带;而籼粳杂合子F1扩增出458bp和562bp(或561bp)的特征条带。所设计引物均选择在基因的外显子区域,保证了引物的适用性和可靠性。经检测验证,以上5条引物在同一体系中进行PCR扩增,能准确区分粳型COLD1Jap、籼型COLD1Ind及其杂合类型。与先前报道的衍生型酶切扩增多态性序列的功能标记相比,该方法省去了内切酶的使用,降低了成本,也省去了酶切时间,避免了使用聚丙烯酰胺凝胶电泳操作复杂等问题,提高了检测效率。同时,利用其中的4条引物进行检测,还弥补了先前不能鉴定籼型COLD1Ind具体变异碱基的不足,是一种简便、高效、快捷的COLD1基因鉴定方法,可用于水稻种质资源COLD1基因籼粳属性的鉴定和分子辅助选择育种,有较好的推广应用价值。
4 结论
根据水稻苗期耐低温基因COLD1在籼粳中的第4外显子存在的SNP2差异,结合扩增受阻突变体系PCR技术原理,设计了用于鉴定该基因的新功能标记。研究结果表明,利用2条正反向外引物和3条正反向内引物组成的5条引物混合在同一反应体系里进行PCR扩增,可准确快速地区分纯合粳型COLD1Jap、纯合籼型COLD1Ind及其杂合基因型。进一步,针对籼型COLD1Ind在SNP2处的两种不同碱基类型,可用2条正反向外引物和各1条正反向内引物组成的4引物混合在同一反应体系里进行PCR扩增鉴定。本研究所设计的新功能标记检测方法操作简便,费用低廉,弥补了先前报道的衍生型酶切扩增多态性序列功能标记存在的费用昂贵、操作复杂及难以对籼型COLD1Ind具体差异碱基类型鉴别等的不足,可广泛应用于水稻COLD1基因的资源鉴定和分子标记辅助选择育种。
参考文献
Identification and mapping of quantitative trait loci controlling cold-tolerance of Chinese common wild rice (O. rufipogon Griff.) at booting to flowering stages
QTLs conferring cold tolerance at the booting stage of rice using recombinant inbred lines from a japonica×indica cross
Identification and mapping of quantitative trait loci for cold tolerance at the booting stage in a japonica rice near-isogenic line
Quantitative trait locus analysis for cold tolerance at the booting stage in a rice cultivar,hatsushizuku
Mapping quantitative trait loci for cold tolerance at the booting stage in rice by using chromosome segment substitution lines
Delimitation of a QTL region controlling cold tolerance at booting stage of a cultivar,'Lijiangxintuanheigu',in rice,Oryza sativa L
Mapping and validation of QTLs for cold tolerance at seedling stage in rice from an indica cultivar Habiganj Boro VI (Hbj. BVI)
Map-based cloning of the rice cold tolerance gene Ctb1
Cold sensitivity in rice (Oryza sativa L) is strongly correlated with a naturally occurring I99V mutation in the multifunctional glutathione transferase isozyme GSTZ2
Molecular identification of a major quantitative trait locus,qLTG3-1,controlling low-temperature germinability in rice
Rice LTG1 is involved in adaptive growth and fitness under low ambient temperature
COLD1 confers chilling tolerance in rice
A novel functional gene associated with cold tolerance at the seedling stage in rice
Natural variation in CTB4a enhances rice adaptation to cold habitats
Early selection of bZIP73 facilitated adaptation of japonica rice to cold climates
Natural variation in the HAN1 gene confers chilling tolerance in rice and allowed adaptation to a temperate climate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}