


开放科学(资源服务)标识码(OSID): 
玉米播种后的田间温度环境是动态变化的,而针对低温冷害对玉米萌发与幼苗生长影响的报道大多是在恒定的适宜温度条件下开展的,难以准确解析玉米幼苗生长及群体建成对田间阶段性低温冷害的响应机制。鉴于此,本研究基于寒地典型玉米种植生态区的气象条件,在播种至苗期分阶段进行人工模拟低温胁迫环境,通过探讨玉米出苗进程、主要农艺性状及相关抗性生理指标的变化差异,明确播种至苗期阶段性低温和低温持续时间对玉米幼苗建成的影响规律,为不断创新寒地玉米全程机械化丰产增效关键技术、改善农田管理措施及防灾避灾能力、科学合理应对气候变化提供理论依据。
1 材料与方法
1.1 试验材料
供试玉米品种为先玉335(由铁岭先锋种子研究有限公司选育),供试土壤取自黑龙江八一农垦大学试验实习基地(大庆)农田0~20cm耕层,土壤类型为草甸土,基本理化性质为:碱解氮104.46mg/kg,速效磷11.47mg/kg,速效钾113.73mg/kg,有机质26.59g/kg,pH值8.06。
1.2 试验设计
试验于2018年在黑龙江省现代农业栽培技术与作物种质改良重点实验室进行,选取长势饱满、大小均匀一致的玉米种子,播于装有1 600g风干土的PVC培养钵(长×宽×高:13.5cm×13.5cm×12.0cm)中,每钵播种16粒,播深4cm。播后采用称重法进行统一均匀灌溉,然后置于温度(-10~45)±0.5℃、湿度(10%~95%)±3%、光量子密度450µmol/(m2·s)的人工气候箱中进行低温模拟试验。根据黑龙江省典型玉米种植生态区2008-2017年的气象资料数据所拟合的春季耕层0~5cm土壤温度日变化趋势(图1),动态模拟播后至苗期玉米生长温度环境,采用二因素完全随机设计,分别设置低温处理阶段和低温持续时间处理。低温处理阶段设置播后第1~5(P1)、6~10(P2)、11~15(P3)、16~20(P4)、21~25(P5)、26~30(P6)天6个阶段,低温持续时间分别设置4℃低温处理0(D0)、1(D1)、2(D2)、3(D3)、4(D4)、5d(D5)6个水平,以0d(常规温度)培养为对照(D0),总计31个处理组合,且均为4次重复,每个培养钵作为1次重复。试验过程中,控制人工气候箱湿度为(60±5)%,光照强度为450µmol/(m2·s),光照时间为12h/d,每隔1d进行统一补水。播种后,每天记录各处理组合幼苗的破土和出苗情况,于播后第35天选取各培养钵具代表性植株6株,检测功能叶片叶绿素含量与叶绿素荧光参数,并对幼苗形态和抗性生理指标进行测定。
图1
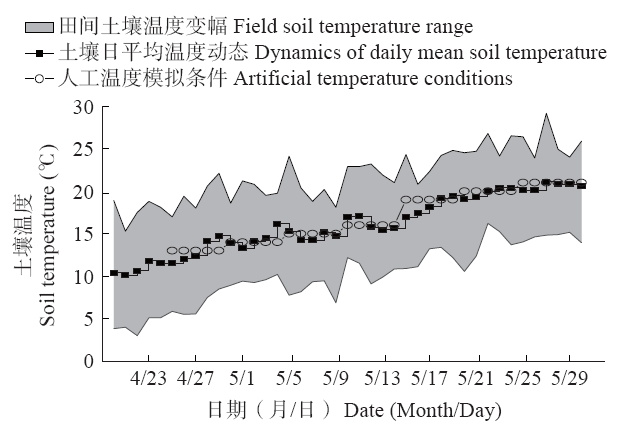
图1
春季日平均地温变化趋势及温度人工模拟条件
Fig.1
Variation trend for the daily average ground temperature in spring and the artificial simulated conditions of temperature
1.3 测定项目与方法
1.3.2 叶片叶绿素含量及叶绿素荧光参数的测定
选定玉米幼苗最新完全展开功能叶片,将叶片暗适应30min后,利用叶绿素脉冲制荧光分析仪OS-30p(Opti-Sciences,美国)分别测定叶片上、中、下3部分的叶绿素荧光参数,分别读取初始荧光Fo,待Fo稳定后照射饱和脉冲光,获得最大荧光Fm,并计算最大光化学效率(Fv/Fm),Fv/Fm=(Fm-Fo)/Fm;于相同位置利用叶绿素仪SPAD-502 Plus(Konica Minolta Sensing,日本)同步测定相对叶绿素含量(SPAD值),最后取平均值[13]。
1.3.3 幼苗形态指标的测定 首先使用自来水将幼苗根部土壤缓慢冲洗干净,用吸水纸吸干表面水分,然后使用直尺测量具代表性植株的株高、初生胚根长、胚芽鞘长度、叶片长度和宽度,用游标卡尺测量茎基部直径;采用长宽系数法(长×宽×0.75)计算叶面积[14]。
1.3.4 植株鲜重、干重及贮藏物质运转率的测定
将取样的植株分成地上部、地下部和残留种子3部分,使用Presica LS120A万分之一天平分别称量鲜重后,置于牛皮纸袋中于105℃杀青30min,然后置于80℃下烘干至恒重后称重,并计算贮藏物质运转率,贮藏物质运转率=幼苗干重/(幼苗干重+残留种子干重)×100%[15]。
1.4 数据处理与分析
采用SPSS 22.0进行数据的统计分析,通过Duncan多重比较和t检验分析不同处理的影响差异,利用多因素方差分析因素间的交互效应,运用Microsoft Excel 2003及Origin 2018绘制图表。
2 结果与分析
2.1 播后不同阶段低温与低温处理时间对玉米出苗进程的影响
2.1.1 出苗速率 随着播后不同阶段低温处理时间的延长,玉米的出苗速率逐渐下降,且各阶段均在D5处理下达最低值(图2)。从表1可以看出,低温处理阶段、低温持续时间单因素对出苗速率的影响均达极显著水平,但二者的交互效应并不明显,其中P4和P5处理下的出苗速率分别较P1、P2、P3和P6下降了5.01%~13.13%、2.73%~11.04%,D0、D1和D2处理下的出苗速率分别较D4和D5高出15.06%和15.88%、7.29%和8.06%、4.94%和5.69%,且均达显著水平。从不同处理组合来看,P4D5处理的出苗速率最低,为0.038,除了与P2D5(0.0409)、P3D5(0.0418)、P4D2(0.0418)、P4D3(0.0400)、P4D4(0.0387)以及P5D2(0.0418)、P5D3(0.0413)、P5D4(0.0396)、P5D5(0.0396)处理差异不明显以外,较其他处理显著降低了11.24%~22.34%。
图2
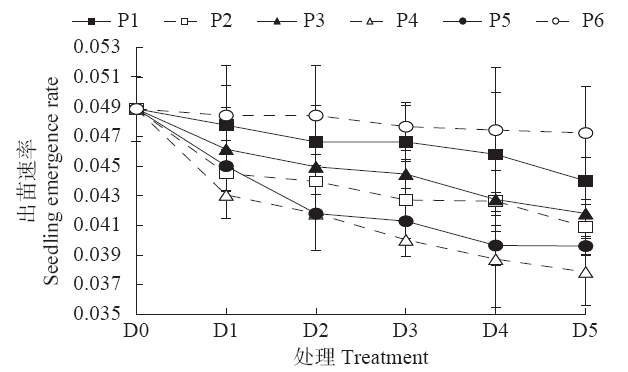
图2
各阶段不同低温持续时间对玉米出苗速率的影响
Fig.2
Effects of different low temperature durations at different stages on seedling emergence rate of maize
表1 不同阶段低温处理、低温持续时间对玉米幼苗出苗速率和成苗率的交互效应
Table 1
| 试验因素 Experimental factor | 处理 Treatment | 出苗速率 Seedling emergence rate | 成苗率 Seedling survival rate (%) |
|---|---|---|---|
| P | P1 | 0.046b | 93.49a |
| P2 | 0.044cd | 93.23a | |
| P3 | 0.045bc | 92.97a | |
| P4 | 0.042e | 89.58a | |
| P5 | 0.043de | 91.41a | |
| P6 | 0.048a | 92.45a | |
| D | D0 | 0.049a | 98.44a |
| D1 | 0.046b | 96.35ab | |
| D2 | 0.045bc | 93.49bc | |
| D3 | 0.044cd | 90.89cd | |
| D4 | 0.043d | 88.54de | |
| D5 | 0.042d | 85.42e | |
| F值F-value | P | 17.44** | 1.42 |
| D | 19.18** | 13.84** | |
| P×D | 1.06 | 0.53 |
注:同列数据后不同字母表示处理间差异达0.05显著水平;*表示P<0.05,**表示P<0.01,下同
Note: Data followed by different letters within a column indicate significant difference at the 0.05 level between treatments;* indicates P < 0.05, ** indicates P < 0.01, the same below
图3
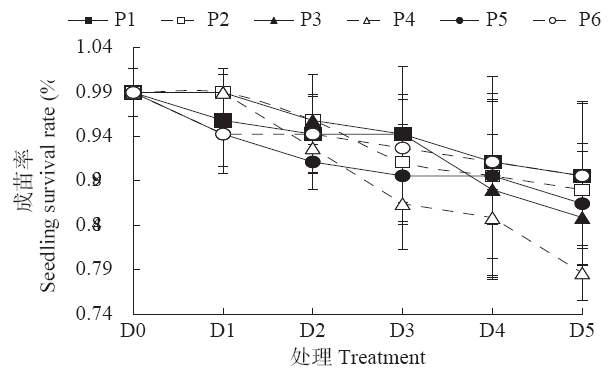
图3
各阶段不同低温持续时间对玉米成苗率的影响
Fig.3
Effects of different low temperature durations at different stages on seedling survival rate of maize
2.2 播后不同阶段低温与低温处理时间对玉米幼苗形态指标的影响
如表2所示,低温处理阶段及低温处理时间单因素对玉米幼苗株高、茎基部直径、叶面积、胚芽鞘长和初生胚根长的影响均达显著(P<0.05)或极显著水平(P<0.01)。P1和P2阶段随着低温处理天数的增加,幼苗株高、茎基部直径变化幅度较小,而在P3~P6阶段,株高和茎基部直径总体上明显受低温处理影响,以P4D5处理的株高和茎基部直径最小,比对照处理分别下降了26.89%和19.54%。同时,相比于对照处理,P1~P6阶段低温处理后的幼苗叶面积和初生胚根长均有所减小,叶面积降幅分别为12.40%~28.11%、17.69%~29.10%、18.94%~29.14%、31.96%~48.07%、22.57%~39.56%和16.29%~34.31%,初生胚根长降幅分别为10.73%~31.35%、18.37%~30.01%、18.65%~37.59%、26.86%~42.15%、21.32%~37.31%和23.07%~38.36%,且均达显著水平;而胚芽鞘长仅在P3D5、P4D3、P4D4和P4D5处理组合下有明显的增长趋势。从两因素互作方面来看,低温处理阶段与低温处理时间仅对幼苗叶面积大小具有极显著的交互效应。
表2 不同阶段低温处理和低温持续时间对玉米幼苗形态指标及生物量积累的影响
Table 2
| 处理 Treatment | 株高 Plant height (cm) | 茎基部直径 Diameter for basal part of stem (cm) | 叶面积 Leaf area (cm2) | 胚芽鞘长 Coleoptile length (cm) | 初生胚根长 Primary radicle length (cm) | 植株鲜重 Plant fresh weight (g) | 植株干重 Plant dry weight (g) | 贮藏物质转运率 Storage substance transportation ratio (%) |
|---|---|---|---|---|---|---|---|---|
| P1D0 | 17.48a | 0.348a | 27.25a | 1.89e | 14.26a | 1.47a | 0.177a | 59.6a |
| P1D1 | 16.89ab | 0.335ab | 23.87b | 1.96bcde | 12.73b | 1.42ab | 0.161ab | 59.0ab |
| P1D2 | 16.62abc | 0.327abcd | 22.15cde | 1.97bcde | 11.77c | 1.36abcde | 0.151abc | 57.5abc |
| P1D3 | 16.48abcd | 0.326abcd | 21.02cdefghi | 1.97bcde | 11.48cd | 1.30abcdef | 0.15abc | 54.3abcd |
| P1D4 | 16.33abcd | 0.326abcd | 19.90ghij | 1.98abcde | 10.63defghi | 1.27abcdef | 0.145abcd | 53.7abcd |
| P1D5 | 16.08bcde | 0.318bcdefgh | 19.59hijk | 1.99abcde | 9.79hijklm | 1.22abcdefgh | 0.138bcde | 52.9abcde |
| P2D0 | 17.48a | 0.348a | 27.25a | 1.89e | 14.26a | 1.47a | 0.177a | 59.6a |
| P2D1 | 16.33abcd | 0.329abc | 22.43bcd | 1.96bcde | 11.64cdef | 1.42ab | 0.163abc | 55.9abc |
| P2D2 | 16.28abcd | 0.325abcde | 21.33cdefgh | 1.97bcde | 11.23cdefg | 1.31abcdef | 0.155abcd | 52.5abcde |
| P2D3 | 16.24abcd | 0.322abcdef | 20.69defghi | 1.98bcde | 10.78fghijk | 1.23abcdefgh | 0.145abcd | 52.3abcde |
| P2D4 | 15.96bcde | 0.321abcdefg | 20.24fghij | 1.98abcde | 10.13hijklm | 1.18bcdefgh | 0.135bcde | 50.7bcdef |
| P2D5 | 15.61bcdefg | 0.320bcdefg | 19.32ijkl | 2.00abcde | 9.98mn | 1.15cdefghi | 0.121bcde | 50.7bcdef |
| P3D0 | 17.48a | 0.348a | 27.25a | 1.89e | 14.26a | 1.47a | 0.177a | 59.6a |
| P3D1 | 16.28abcd | 0.302cdefghi | 22.09cdef | 2.01abcde | 11.6cd | 1.38abcd | 0.151ab | 56.7abc |
| P3D2 | 15.97bcde | 0.302cdefghi | 21.07cdefghi | 2.02abcde | 11.31cde | 1.24abcdefg | 0.147abcd | 52.9abcde |
| P3D3 | 15.78bcdef | 0.290hi | 20.41efghi | 2.04abcde | 10.91cdefgh | 1.20bcdefgh | 0.143bcde | 51.2abcdef |
| P3D4 | 15.71bcdefg | 0.290hi | 19.89ghij | 2.08abcde | 9.96fghijk | 1.15cdefghi | 0.139bcde | 50.1cdef |
| P3D5 | 15.27defg | 0.288i | 19.31ijkl | 2.10abcd | 8.90fghijk | 1.12defghi | 0.136bcdef | 49.3cdef |
| P4D0 | 17.48a | 0.348a | 27.25a | 1.89e | 14.26a | 1.47a | 0.177a | 59.6a |
| P4D1 | 14.90efgh | 0.296fghi | 18.54jkl | 2.05abcde | 10.43efghij | 1.21bcdefgh | 0.147abcd | 54.6abcd |
| P4D2 | 13.90hijk | 0.293ghi | 16.81mn | 2.09abcde | 9.90ghijkl | 1.05fghij | 0.138bcde | 50.1cdef |
| P4D3 | 13.43ijk | 0.290i | 15.60nop | 2.11abc | 9.68ijklm | 0.98hijk | 0.122cdef | 45.8defg |
| P4D4 | 13.27jk | 0.288i | 15.05op | 2.13ab | 9.32klm | 0.87jk | 0.106ef | 44.6efg |
| P4D5 | 12.78k | 0.280i | 14.15p | 2.19a | 8.25n | 0.79k | 0.099f | 39.5g |
| P5D0 | 17.48a | 0.348a | 27.25a | 1.89e | 14.26a | 1.47a | 0.177a | 59.6a |
| P5D1 | 16.09bcde | 0.324abcdef | 21.10cdefghi | 1.90cde | 11.22cde | 1.29abcdef | 0.163ab | 57.0abc |
| P5D2 | 15.54cdefg | 0.307bcdefghi | 20.69defghi | 1.91cde | 9.85hijkl | 1.24abcdefg | 0.144abcd | 56.6abc |
| P5D3 | 14.47ghij | 0.305cdefghi | 19.93ghij | 1.92bcde | 9.47jklm | 1.17bcdefgh | 0.137bcde | 49.3cdef |
| P5D4 | 13.84hijk | 0.303cdefghi | 17.67lm | 1.93bcde | 9.29klm | 0.99ghijk | 0.114def | 44.3efg |
| P5D5 | 13.65ijk | 0.300defgji | 16.47mno | 1.94bcde | 8.94lmn | 0.92ijk | 0.113def | 43.3fg |
| P6D0 | 17.48a | 0.348a | 27.25a | 1.89e | 14.26a | 1.47a | 0.177a | 59.6a |
| P6D1 | 15.51cdefg | 0.321abcdef | 22.81bc | 1.92de | 10.97cd | 1.38abc | 0.161ab | 57.0abc |
| P6D2 | 15.42cdefg | 0.318bcdefgh | 21.63cdefg | 1.88bcde | 10.87cde | 1.29abcdef | 0.143abc | 53.9abcd |
| P6D3 | 14.89efgh | 0.297efghi | 21.04cdefghi | 1.93bcde | 10.14cdef | 1.15cdefghi | 0.137abcd | 52.1abcde |
| P6D4 | 14.55fghi | 0.297efghi | 19.67hijk | 1.93bcde | 9.78fghijk | 1.15cdefghi | 0.133bcde | 51.8abcdef |
| P6D5 | 14.48ghij | 0.290i | 17.90klm | 1.95bcde | 8.79lmn | 1.12efghi | 0.128cdef | 49.9cdef |
| F值F-value | ||||||||
| P | 19.76** | 13.89** | 50.70** | 6.16** | 18.77** | 5.89** | 2.74 | 3.31** |
| D | 27.20** | 27.67** | 216.98** | 3.55* | 200.51** | 15.04** | 17.73** | 12.17** |
| P×D | 1.21 | 0.80 | 2.71** | 0.31 | 1.48 | 0.39 | 0.39 | 0.46 |
2.3 播后不同阶段低温与低温处理时间对玉米幼苗生物量的影响
低温处理阶段、低温处理时间单因素对玉米幼苗植株鲜重和贮藏物质转运率的影响达极显著水平(P<0.01),植株干重仅受低温处理时间的显著影响(P<0.05)(表2)。尽管播后不同阶段植株鲜重、干重和贮藏物质转运率总体表现为D0>D1>D2>D3>D4>D5,但各阶段不同低温处理时间的影响程度存在差异,其中植株鲜重在P2阶段D4~D5处理、P3阶段D3~D5处理、P4阶段D2~D5处理、P5阶段D3~D5处理、P6阶段D4~D5处理比对照处理分别显著(P<0.05)降低了0.291~0.316、0.271~0.349、0.420~0.680、0.296~0.547和0.322~0.354g,植株干重分别显著降低了0.042~0.056g、0.034~0.041g、0.039~0.078g、0.040~0.064g和0.044~0.049g;此外,种子贮藏物质转运率以P4D5处理最低,除与P4D3、P4D4、P5D4和P5D5差异不明显外,显著低于其他处理19.88%~33.72%。
2.4 播后不同阶段低温与低温处理时间对叶片相对叶绿素含量及叶绿素荧光参数的影响
从图4和表3可以看出,低温处理阶段及低温持续时间单因素对玉米幼苗叶片SPAD值和Fv/Fm均具有极显著(P<0.01)影响,且低温处理阶段与低温持续时间处理对叶片SPAD值的影响存在显著(P<0.05)的交互效应。随着低温处理时间的延长,叶片SPAD值和Fv/Fm表现为逐渐下降的趋势,而低温处理阶段单因素影响总体表现为P1>P2>P3>P6>P5>P4,其中在P1和P2阶段D3~D5处理、P3阶段D2~D5处理下叶片SPAD值明显低于对照处理,而P4~P6阶段各低温持续时间处理均显著下降,其中P4D4和P4D5处理组合的SPAD值相对较低,分别为26.79和25.79。而在P4~P6阶段低温处理影响下的Fv/Fm降幅为4.17%~6.65%。
图4

图4
不同阶段低温处理及低温持续时间条件下叶片叶绿素含量及最大光化学效率
Fig.4
Leaf chlorophyll content and maximum photochemical efficiency of maize seedlings effected by low temperature with different phases and various durations
表3 不同阶段低温、低温持续天数对玉米幼苗光合作用与抗性生理特性的交互效应
Table 3
| 试验因素 Experimental factor | 处理 Treatment | SPAD | Fv/Fm | 可溶性蛋白含量 Soluble protein content (mg/g FW) | 脯氨酸含量 Proline content (μg/g FW) | 可溶性糖含量 Soluble sugar content (mg/g FW) | 丙二醛含量 Malondialdehyde content (μmol/g FW) |
|---|---|---|---|---|---|---|---|
| P | P1 | 33.94a | 0.764a | 2.85e | 24.90d | 0.62e | 3.45e |
| P2 | 33.72a | 0.760a | 2.98d | 25.28cd | 0.67d | 3.66d | |
| P3 | 33.35a | 0.752b | 3.00d | 26.05c | 0.80c | 4.11c | |
| P4 | 29.74c | 0.737d | 3.33a | 30.52a | 0.93a | 4.55a | |
| P5 | 31.30b | 0.743cd | 3.29b | 29.11b | 0.83b | 4.35b | |
| P6 | 31.92b | 0.749bc | 3.11c | 28.74b | 0.81c | 4.14c | |
| D | D0 | 36.13a | 0.767a | 2.37f | 20.66f | 0.44f | 2.95f |
| D1 | 33.35b | 0.756b | 2.73e | 23.24e | 0.62e | 3.71e | |
| D2 | 32.55b | 0.753bc | 2.96d | 25.82d | 0.72d | 4.01d | |
| D3 | 31.33c | 0.747cd | 3.23c | 29.38c | 0.83c | 4.26c | |
| D4 | 30.71cd | 0.742de | 3.52b | 31.36b | 0.94b | 4.51b | |
| D5 | 29.91d | 0.738e | 3.75a | 34.16a | 1.11a | 4.81a | |
| F值F-value | P | 29.28** | 14.17** | 382.70** | 72.26** | 450.61** | 188.36** |
| D | 54.20** | 15.42** | 2 921.27** | 349.68** | 1 940.62** | 470.94** | |
| P×D | 1.80* | 0.93 | 86.20** | 4.97** | 31.30** | 9.32** |
2.5 播后不同阶段低温与低温处理时间对玉米幼苗抗性生理指标的影响
随着玉米播后至苗期低温处理阶段的推迟,以及低温持续时间的延长,叶片可溶性蛋白、可溶性糖、脯氨酸和丙二醛含量呈逐渐增加趋势(图5),且可溶性蛋白、可溶性糖、脯氨酸和丙二醛含量均受低温处理阶段、低温处理天数单因素的极显著(P<0.01)影响,同时两因素的互作效应亦达极显著水平(表3)。其中,叶片可溶性蛋白、可溶性糖、脯氨酸和丙二醛含量在D5处理时达到峰值,分别较D0处理高57.99%、149.44%、65.32%和62.97%;低温处理阶段单因素的影响总体表现为P4>P5>P6>P3>P2>P1,P4和P5阶段的叶片可溶性蛋白、可溶性糖、脯氨酸和丙二醛含量较其他处理分别增加了0.17~0.47mg/g、0.02~0.31mg/g、0.36~5.61µg/g和0.21~1.10μmol/g,且均达显著水平。
图5

图5
不同阶段低温处理及低温持续时间条件下叶片抗性生理指标的变化
Fig.5
Changes of leaf physiological indexes for resistance effected by low temperature with different phases and various durations
3 讨论
东北地区,特别是黑龙江省玉米生产中常发生春季低温冷害,影响玉米高产群体建成和产量稳步提升。玉米种子的萌发不仅需要足够的水分和充足的氧气环境,适宜的温度条件也至关重要。低温逆境除了抑制玉米种子的萌发进程,还会影响出苗整齐度,降低成苗率[19]。本试验中,P1阶段1~4d的低温处理对出苗速率影响不显著,说明在种子物理吸涨过程中遇到低温逆境对玉米出苗速率影响较小,但在破土前后的P3和P4阶段,低温对出苗速率的影响非常明显,而P6阶段由于已完成出苗,出苗速率未受到低温处理的影响。同时,本试验中玉米成苗率主要随低温持续时间的增加呈显著下降趋势,且仅在P3~P4阶段经历长时间(3~5d)的持续低温处理后成苗率显著降低,这可能是由于长期低温处理导致植物体内能量的亏缺导致植株的死亡[20],同时张海艳[21]研究发现植株地上部比地下部对温度响应更为敏感,严重抑制了破土至出苗临界期玉米胚根和胚芽的快速生长,甚至导致植株生长停滞和死亡。此外,随着玉米幼苗生育进程的推进,尽管P5和P6阶段耐冷性有所提高,但在长时间持续低温条件下成苗率也会有小幅度下降的趋势。
已有研究表明,低温胁迫后玉米幼苗生长将受到明显抑制,初生胚根长、株高、茎基部直径、地上部鲜重、根系鲜重及叶面积出现明显的下降趋势[22],胚芽鞘的相对生长率也显著高于正常生长植株[23]。本研究结果表明,播后各阶段随着低温处理时间的延迟,玉米幼苗的株高、茎基部直径、叶面积、初生胚根长、生物量积累呈逐渐降低趋势,相对而言,以P3~P6阶段受低温处理的影响较为明显;尽管胚芽鞘长随低温持续时间的增加呈逐渐增长趋势,但仅在P3D5、P4D3、P4D4和P4D5处理组合下影响达显著水平。刘禹辰等[24]认为种子萌发的过程是淀粉等贮藏物质水解为小分子物质的过程,水通过促进种子吸胀,引起种子内各种酶活性增强,降解种子贮藏物质,促进种子萌发。本研究中,相比于对照处理,P2和P3阶段4~5d、P4阶段2~5d、P5阶段3~5d和P6阶段5d的低温处理明显抑制了贮藏物质转运率,这可能是持续低温影响种子呼吸代谢途径及幼苗碳源与能量供应的结果[25]。
植物细胞中可溶性蛋白、脯氨酸以及可溶性糖是重要的渗透调节物质,对维持细胞膨压、缓解低温胁迫对细胞造成的损伤具有重要作用[26],而丙二醛是植物在逆境胁迫下膜脂过氧化的分解产物,其含量的高低可以有效反映细胞受伤害的程度[27]。Hendrickson等[28]和田礼欣等[29]研究发现,低温胁迫可降低光合色素的合成速率,加速叶绿素的降解,引起膜脂过氧化,抑制光能的吸收与传递,最终致使光合速率降低。本试验条件下,随低温持续时间的延长,幼苗叶片可溶性蛋白、可溶性糖、脯氨酸与丙二醛含量在各阶段均呈增加趋势,而SPAD值和Fv/Fm逐渐降低;随着低温处理阶段的延后,渗透调节物质与丙二醛含量呈先增加后下降的趋势,而SPAD值和Fv/Fm呈先降低后升高趋势,分别在P4阶段达极值,这进一步说明,尽管播种后至苗期各阶段的低温处理均会引起幼苗光合作用效率及抗性生理特征的响应,但这种低温胁迫的影响程度因低温处理时期的不同而有所差异,以P3~P5阶段受到影响较为明显,特别是长时间持续低温将对植株造成一系列不可逆的生理损伤,从而引起成苗率的显著下降。
4 结论
播种后至苗期不同阶段的低温胁迫,可不同程度地延长玉米出苗时间,降低成苗率,抑制玉米幼苗的生长及生物量积累,引起玉米幼苗叶片SPAD值和Fv/Fm的降低,进而引进渗透调节物质含量的增加与膜脂过氧化程度的加剧,促使幼苗素质降低,弱苗比例增加,难以构建高产群体;这些不利影响的程度随低温持续时间的延长而不断加剧,且以播种后11~25d,特别是16~20d,即处于破土至出苗临界期的玉米植株对低温胁迫最为敏感。
参考文献
水分和温度对春玉米出苗速度和出苗率的影响
DOI:10.5846/stxb201105050585
URL
[本文引用: 1]

Zea mays)播种出苗期间土壤含水量与出苗期和出苗率的关系均呈显著的二次函数关系。在田间持水量以下,耕层土壤湿度越大,玉米出苗越快,出苗率越高;干旱推迟出苗期,降低出苗率。气温低出苗慢,但温度对出苗率无明显影响。播种出苗期间0-20 cm深平均土壤湿度(S)、土壤有效水量(H)和平均气温(T)对出苗速度的组合影响指标是:25%>S>21%(或70 mm>H>45 mm)、T>18 ℃ 玉米出苗快;21%>S>18%(或45 mm>H>35 mm)、18 ℃>T>16.5 ℃ 玉米出苗比较快;18%>S>16%(或35 mm>H>25 mm)、16.5 ℃>T>15.5 ℃玉米出苗较慢;SHTH>50 mm)出苗率最高(95%以上);相对湿度80%左右(或50>H>40 mm)出苗率较高(90% 左右);相对湿度70%以下(或HH
5-氨基乙酰丙酸提高玉米幼苗抗低温性及其生理机制
低温是影响东北地区玉米生长发育的主要因素之一。5-氨基乙酰丙酸(ALA)具有增强作物抗逆性的作用,近年来成为国内外的研究重点。为了探究ALA缓解玉米幼苗低温胁迫的生理机制及叶面喷施的最适浓度,选取“绥玉13号”为试验材料,叶面喷施不同浓度的ALA(0、10、20、30和40 mg·L-1),在昼/夜温度为14 ℃/5 ℃的低温条件下培养48 h,研究ALA对玉米幼苗生长、细胞膜稳定性、抗氧化酶活性及渗透调节物质的影响。结果表明:叶面喷施ALA能促进低温胁迫下幼苗生长,保护叶片细胞膜的结构与功能,增强细胞清除活性氧能力,提高渗透调节的能力;其中以喷施20 mg·L-1 ALA效果最佳,与对照相比均达到显著水平(P<0.05),幼苗株高、叶面积和鲜(干)重分别增加24%、23%和26%(24%),MDA含量和相对电导率减少38%和45%,SOD和POD活性增加37%和66%,可溶性蛋白和可溶性糖含量增加50%和45%;叶面喷施适宜浓度的ALA可以缓解低温胁迫对玉米幼苗的损伤。 ]]>
温度对玉米生长和产量的影响
为了明确气温变化对玉米生长发育和产量的影响,建立玉米低温冷害监测和气象评估模式,在东北地区中部的榆树市进行了玉米分期播种试验。试验采用3个品种,设置早、中、晚3个播种期,进行玉米生长发育进程、叶面积指数、生物量、产量和温度等观测。结果表明:玉米的生长速率与温度密切相关,平均气温每升高1 ℃,出苗速率提升17%,营养生长速率提升5%;积温每增加100 ℃·d,玉米最大叶面积指数增加10%,最大生物量和产量增加8%;玉米生长发育期(日平均气温稳定>10 ℃期间)间平均气温降低0.7 ℃,或活动积温减少100 ℃·d,玉米成熟期将延迟7 d,发生一般低温冷害,玉米单产减少8%;气温下降1 ℃,或积温减少140 ℃·d,生育期延迟10 d,发生严重低温冷害,减产10%以上;在水分条件比较适宜的前提条件下,气候变暖对东北地区玉米单产提高是有利的。
种子引发对盐胁迫下高粱种子萌发及幼苗生长的影响
-1?NaCl 溶液对种子进行引发处理。采用营养液沙培试验, 设4 个NaCl 浓度(0、50 mmol?L-1、100 mmol?L-1和150 mmol?L-1)模拟盐胁迫, 研究种子引发处理对盐胁迫下高粱种子萌发及幼苗生长的影响。结果表明: 随着盐胁迫强度增加, 无论经引发处理还是未经引发处理2 个高粱杂交组合种子的出苗时间均明显延迟, 出苗率和成苗率下降, 幼苗生长受到抑制, 物质积累减少。同等盐胁迫强度下,引发处理与未引发处理相比, 种子出苗时间缩短, 出苗率和成苗率提高, 幼苗地上部分干、鲜重和地下部分干、鲜重增加, 光合色素含量升高, Na+/K+值显著降低。由此得出, 种子引发处理可以不同程度地促进盐胁迫下高粱种子萌发, 减轻盐胁迫对幼苗的伤害, 促进幼苗生长, 提高耐盐性; 耐盐性不同的品种引发效果存在差异, 对耐盐性较弱的品种引发效果更好。]]>
外源糖调控玉米光合系统和活性氧代谢缓解盐胁迫
DOI:10.3864/j.issn.0578-1752.2014.20.004
URL
[本文引用: 1]
0.5 mmol·L-1 Glc和Suc预处理3 d,以1/2 Hoagland营养液为空白对照,以等浓度甘露醇为渗透对照;然后,采用150 mmol·L-1 NaCl(1/2 Hoagland营养液配制)对玉米幼苗进行盐胁迫处理4—6 d,以相应糖预处理和空白处理玉米的非盐胁迫为对照。于处理第4天取第2片全展叶进行玉米光合相关和抗氧化等各项生理生化指标测定;于处理第6天进行株高、根长和干重测定;同时于处理第2、4、6天采用Li-6400XTR光合仪测定光合参数的动态变化。【结果】与空白未处理对照相比,Glc和Suc预处理导致玉米幼苗根长、干重和叶片含水量增加,但是显著提高玉米幼苗株高,分别增高了6.39%和4.86%,并维持叶片较高的净光合速率(Pn),其它供试生理生化指标变化不显著;同时,盐胁迫可明显抑制玉米幼苗生长,盐处理6 d株高和根长降低了16.37%和12.17%,地上部及地下部干重分别降低了20.69%和25.00%。盐处理导致玉米叶片中硫代巴比妥酸反应物(TBARS)和过氧化氢(H2O2)含量显著增多,抗氧化酶的活性显著降低,叶片Pn、气孔导度(Gs)、蒸腾速率(Tr)、Hill反应活性, 原初光能转换效率(Fv/Fm)、光合色素含量显著下降, 其中胞间CO2浓度(Ci)随着胁迫时间的延长呈先下降后上升的趋势;重要的是, 外源Glc和Suc预处理可显著促进盐胁迫下玉米幼苗生长和干物质积累, 其中Glc预处理的玉米株高、根长、地上和地下干重较S处理分别增加6.12%、4.49%、15.65%、18.52%, Suc预处理分别增加7.34%、4.87%、17.39%、22.22%。外源Glc和Suc预处理能有效改善玉米幼苗的光合作用, 显著减缓盐胁迫下玉米叶片Pn、Tr、Gs、Fv/Fm的下降速度, 缓解Ci的升降速度,提高叶片Hill反应活性,增加叶片持水量和光合色素的含量。同时,Glc和Suc预处理还显著减少盐胁迫下玉米叶片中硫代巴比妥酸反应物(TBARS)和过氧化氢(H2O2)的含量,提高叶片超氧化物歧化酶(SOD)、抗坏血酸过氧化物酶(APX)、谷胱甘肽过氧化物酶(GPX)的活性,其中,Suc预处理在清除H2O2及提高叶片Hill反应活性两方面均显著高于Glc。此外,与空白盐胁迫处理相比,除引起玉米叶片Hill反应和叶绿素a(Chl a)的显著变化外,采用等渗的甘露醇预处理玉米幼苗其供试生理生化指标未发生显著改善,亦未能引起供试生物学指标的显著改善。【结论】正常生长条件下,Glc和Suc处理可通过维持叶片较高的Pn,促进玉米幼苗地上部分生长;盐胁迫条件下,外源Glc和Suc不仅维持玉米幼苗较高Pn,而且通过增强光合膜系统稳定性、增加光合色素含量、提高PSII反应活性、改善玉米叶片气孔开放及碳同化能力等生理生化机制提高玉米幼苗的耐盐性。]]>
播种深度对夏玉米幼苗性状和根系特性的影响
选用郑单958(ZD958)和先玉335(XY335)为试验材料,在砂培和大田条件下设置3、5、7和9 cm 4个播种深度,并在大田条件下以不同播种深度混播作为对照(CK),研究播种深度对夏玉米幼苗性状和根系特性的影响.结果表明: 随着播种深度的增加,夏玉米的出苗率下降,出苗时间延长.ZD958和XY335播深9 cm的出苗率较3 cm的分别降低9.4%和11.8%,出苗时间较3 cm的均延长1.5 d.随着播种深度的增加,幼苗长度及幼苗整齐度显著降低,中胚轴长度显著增加,胚芽鞘长度差异不显著;初生胚根长度逐渐减小,次生胚根总长度逐渐增加,总根长度差异不显著;幼苗与中胚轴的总干质量增加,总根干质量差异不显著.随播种深度的增加,种子萌发时幼苗各部位可溶性糖含量增加,营养物质消耗量增加,幼苗根系生长速度增加,根系活力降低,总节根数及节根层数增加.播深增加后出苗率及幼苗活力的降低导致收获穗数的显著降低,最终影响产量形成.此外,播深一致有利于群体整齐度的提高和群体性状的改善,从而提高产量.
Growth and development of maize (Zea mays L.) seedlings under chilling conditions in the field
DOI:10.1016/S1161-0301(96)02007-2 URL [本文引用: 1]
Sugar signalling and gene expression in relation to carbohydrate metabolism under abiotic stresses in plants
DOI:10.1007/BF02703574
URL
PMID:16388148
[本文引用: 1]
Sucrose is required for plant growth and development. The sugar status of plant cells is sensed by sensor proteins. The signal generated by signal transduction cascades, which could involve mitogen-activated protein kinases, protein phosphatases, Ca 2+ and calmodulins, results in appropriate gene expression. A variety of genes are either induced or repressed depending upon the status of soluble sugars. Abiotic stresses to plants result in major alterations in sugar status and hence affect the expression of various genes by down- and up-regulating their expression. Hexokinase-dependent and hexokinase-independent pathways are involved in sugar sensing. Sucrose also acts as a signal molecule as it affects the activity of a proton-sucrose symporter. The sucrose trans-porter acts as a sucrose sensor and is involved in phloem loading. Fructokinase may represent an additional sensor that bypasses hexokinase phosphorylation especially when sucrose synthase is dominant. Mutants isolated on the basis of response of germination and seedling growth to sugars and reporter-based screening protocols are being used to study the response of altered sugar status on gene expression. Common cis-acting elements in sugar signalling pathways have been identified. Transgenic plants with elevated levels of sugars/sugar alcohols like fructans, raffinose series oligosaccharides, trehalose and mannitol are tolerant to different stresses but have usually impaired growth. Efforts need to be made to have transgenic plants in which abiotic stress responsive genes are expressed only at the time of adverse environmental conditions instead of being constitutively synthesized.
玉米幼苗抗氧化酶活性、脯氨酸含量变化及与其耐寒性的关系
研究了黄C(耐寒型)和Mo17(低温敏感型)两种耐寒性不同的玉米自交系幼苗在低温逆境(5 ℃,3 d)下,根、中胚轴和胚芽鞘3个部位过氧化氢酶(CAT)、过氧化物酶(POD)活性和脯氨酸(Pro)含量变化及与其耐寒性的关系.结果表明,低温期间,黄C的中胚轴、胚芽鞘相对生长率显著高于Mo17;黄C的根、中胚轴、胚芽鞘3个部位的CAT、POD活性和Pro含量变化率(其中,中胚轴分别为39.02%、-31.35%和86.86%)均高于Mo17(中胚轴变化率分别为1.86%、-48.67%和12.01%);黄C 3个部位的质膜透性(中胚轴为22.05%)均低于Mo17(中胚轴为29.11%).逐步回归分析表明黄C低温期间积累的干物质主要与Pro含量有关,而Mo17主要与POD活性有关,玉米幼苗中胚轴对低温反应最敏感,中胚轴Pro含量变化率与玉米耐寒性关系最密切.]]>
Sugars,signalling,and plant development
DOI:10.1093/jxb/err379
URL
PMID:22140246
[本文引用: 1]
Like all organisms, plants require energy for growth. They achieve this by absorbing light and fixing it into a usable, chemical form via photosynthesis. The resulting carbohydrate (sugar) energy is then utilized as substrates for growth, or stored as reserves. It is therefore not surprising that modulation of carbohydrate metabolism can have profound effects on plant growth, particularly cell division and expansion. However, recent studies on mutants such as stimpy or ramosa3 have also suggested that sugars can act as signalling molecules that control distinct aspects of plant development. This review will focus on these more specific roles of sugars in development, and will concentrate on two major areas: (i) cross-talk between sugar and hormonal signalling; and (ii) potential direct developmental effects of sugars. In the latter, developmental mutant phenotypes that are modulated by sugars as well as a putative role for trehalose-6-phosphate in inflorescence development are discussed. Because plant growth and development are plastic, and are greatly affected by environmental and nutritional conditions, the distinction between purely metabolic and specific developmental effects is somewhat blurred, but the focus will be on clear examples where sugar-related processes or molecules have been linked to known developmental mechanisms.
Low temperature effects on photosynthesis and growth of grapevine

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}