


TIR(toll-like receptor)基因是TIR-NB-LRR类型的抗病基因(resistance gene,R gene)。TIR蛋白的N端含有白细胞介素受体TIR结构域,能够特异识别病原菌分泌的效应蛋白,协助蛋白质之间的结合[4-5]。目前,已经从烟草[6]、拟南芥[7]和亚麻[8]中鉴定到了TIR基因,并对其功能进行了分析。在烟草中,将TIR蛋白中与Toll或TIR信号转导相关的保守氨基酸位点突变后,突变的转基因烟草植株对烟草花叶病毒的抗性减弱,且病毒可以在突变位点及周围扩散[9]。此外,AtTIR基因突变后,拟南芥植株在没有病原菌诱导情况下的存活率显著降低[10]。同样,在亚麻中发现,TIR结构域突变后,植株的抗病性也显著降低,证实TIR基因在植物抗病性中具有重要作用[11]。迄今为止,对TIR基因的研究主要集中在信号转导及病原菌识别方面。然而,关于苦荞TIR基因克隆与功能等方面的研究还鲜见报道。
苦荞是蓼科(Polygonaceae)荞麦属(Fagopyrum)的一种食药同源的杂粮作物,富含蛋白质、维生素、矿物质及各种药用类黄酮[12]。近年来,随着我国荞麦种植面积的不断扩大,由立枯丝核菌(Rhizoctonia solani)侵害引起的荞麦立枯病害也呈逐年加重趋势,严重阻碍了我国荞麦产业的高质量发展。荞麦立枯病主要发生在苗期,立枯丝核菌通过侵入荞麦种子的种皮或幼苗茎基部和根部的伤口来感染荞麦植株,被感染的荞麦种皮表现为黄色或者褐色,严重时会导致种子腐烂[13]。受到感染的幼苗,前期茎基部会出现暗褐色的病斑或者水泽状,接着病斑部位不断扩大,病斑部位组织凹陷和腐烂,植株发生倒伏现象,后期幼苗发生枯萎,严重时幼苗死亡[14]。目前,荞麦立枯病的防治主要依赖喷施化学药物,虽然能有效杀死致病菌或抑制病菌的生长,但长期使用化学药物会导致土壤微生物种群减少、土壤板结和肥力下降等问题[15]。
为了解析荞麦立枯病菌的致病机理,挖掘抗病基因,本研究从苦荞中克隆了FtTIR基因,对其进行生物信息学分析和基因表达模式分析,并进一步在拟南芥中过表达FtTIR基因进行抗病性鉴定,为研究苦荞TIR基因在立枯丝核菌致病机制提供重要依据。
1 材料与方法
1.1 试验材料及处理方法
供试的立枯丝核菌株、野生型拟南芥(Col-0)、苦荞品种“品苦1号”和植物表达载体pBI121均由中国农业科学院作物科学研究所荞麦基因资源创新研究组提供。
选取颗粒饱满、大小均一的苦荞种子,经蒸馏水浸泡12h后剥去外表皮。依次用1%NaClO和75%乙醇消毒6min,然后用无菌水清洗3~5次,将其平铺于MS培养基上,在22℃、16h/8h光暗周期的培养室中培养。将培养2周后的幼苗用活化好的立枯丝核菌菌柄侵染,菌柄直径约3mm,用菌层接触无菌苗。于侵染后0、6、12、24和48h分别对幼苗根、茎和叶取样,用液氮速冻。
1.2 苦荞总RNA提取和cDNA合成
取苦荞幼苗80~100mg,按照RNA Easy Fast Plant Tissue Kit(北京天根生化科技有限公司)植物组织RNA提取试剂盒提取样品总RNA;用HiScript® III 1st Strand cDNA Synthesis Kit(+gDNAwiper)试剂盒(南京诺唯赞生物技术有限公司)反转录合成第1链cDNA,并于-20℃保存备用。
1.3 FtTIR基因克隆及生物信息学分析
利用CE软件以FtPinG0404132400.01.T01(Pinku1:
表1 引物序列汇总
Table 1
| 引物Primer | 引物序列(5′-3′)Primer sequence (5′-3′) | 用途Function |
|---|---|---|
| FtTIR-F | ATGCAACGTCCAGTAGCCAAA | 基因克隆 |
| FtTIR-R | TTATTTGCTGTTTTCTTCAACCTCG | |
| FtTIR-qPCR-F | GCCTTCTTGCTAATGGG | qRT-PCR |
| FtTIR-qPCR-R | GCTTACAACAACACCTCC | |
| AtPR1-qPCR-F | TCATGGCTAAGTTTGCTTCC | |
| AtPR1-qPCR-R | AATACACACGATTTAGCACC | |
| FtH3-qPCR-F | GAAATTCGCAAGTACCAGAAGAG | qRT-PCR内参基因 |
| FtH3-qPCR-R | CCAACAAGGTATGCCTCAGC | |
| Atactin7-qPCR-F | TCCATGAAACAACTTACAACTCCATCA | |
| Atactin7-qPCR-R | CATCGTACTCACTCTTTGAAATCCACA | |
| 121-FtTIR-F | GAGAACACGGGGGACTCTAGAATGCAACGTCCAGTAGCCAAA | 构建过表达载体 |
| 121-FtTIR-R | CATACCTCCTCCTCCGGATCCTTTGCTGTTTTCTTCAACCTCG | |
| 121F | TGACGTAAGGGATGACGCAC | pBI121通用引物 |
利用NCBI在线网站对测序正确的序列进行blast比对分析,确定FtTIR的开放阅读框(ORF)。利用Expasy(
1.4 立枯丝核菌侵染下苦荞FtTIR基因的表达模式分析
利用CE软件设计FtTIR基因荧光定量PCR引物FtTIR-qPCR-F/R(表1),以苦荞组成型表达基因FtH3为内参基因。使用Taq Pro Universal SYBR qPCR Master Mix荧光定量试剂盒(南京诺唯赞生物技术有限公司)和7500 Real Time PCR System仪器进行qRT-PCR分析。按公式2-ΔΔCT计算基因的相对表达量。
1.5 pBI121-FtTIR过表达载体的构建及拟南芥的遗传转化
以测序比对正确的FtTIR-T质粒为模板,参照pBI121图谱设计特异性引物121-FtTIR-F/R(表1)进行PCR扩增,利用同源重组方法将FtTIR基因的全长编码区连接到空载体pBI121中,测序比对后得到过表达载体质粒pBI121-FtTIR。
将重组质粒pBI121-FtTIR转化到农杆菌GV3101感受态中,筛选鉴定得到阳性克隆。将阳性菌液培养至OD600=0.6~0.8,通过浸花法侵染野生型拟南芥,收取T0代转基因拟南芥种子,然后经卡那霉素抗性筛选得到T1代过表达拟南芥。以T1代过表达拟南芥叶片的DNA为模板,载体pBI121的通用引物121F为正向引物、121-FtTIR-R为反向引物进行PCR以鉴定阳性转基因拟南芥植株。筛选至纯化的T3代转基因拟南芥(FtTIR-OE)并进行后续的生理生化分析。
1.6 pBI121-FtTIR过表达拟南芥的抗病性鉴定
采用离体叶片接种法检测转基因拟南芥的抗病性,将过表达FtTIR拟南芥T3代和野生型拟南芥种子消毒灭菌后先在MS固体培养基上培养,7d后转移到蛭石:营养土=1:1的混合土中培养,培养条件为25℃,光周期为光照16h、黑暗8h。取3周龄大小一致的叶片用于立枯丝核菌侵染。把叶片放置在铺有湿润滤纸的培养皿中,将活化好的立枯丝核菌平板菌打成直径0.3mm的菌饼,菌层接触叶片正面。每个株系取15个叶片,设置3次生物学重复,对照使用无菌PDA培养基。在接菌后24h统计发病情况。进一步做DAB染色试验,并观察其病斑情况,然后检测立枯丝核菌侵染和未侵染条件下拟南芥中FtTIR和病程相关基因AtPR1的相对表达量,并测定过氧化物酶(POD)和超氧化物歧化酶(SOD)活性。
2 结果与分析
2.1 苦荞FtTIR基因的克隆与生物信息学分析
以苦荞幼苗cDNA为模板,经PCR扩增获得一条长度为576bp的目的片段(图1)。
图1
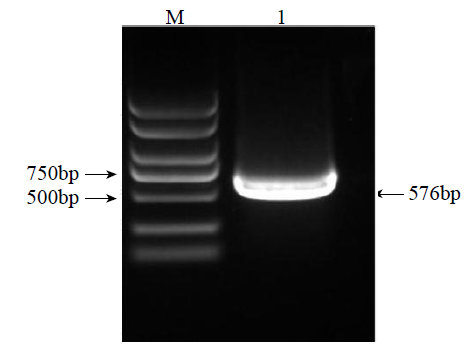
图1
苦荞FtTIR基因序列扩增
M:DL2000;1:FtTIR基因的PCR扩增目的条带
Fig.1
Sequence amplification of FtTIR gene intartary buckwheat
M: DL2000; 1: the PCR amplification target band of FtTIR gene
基因结构分析(图2a)表明,FtTIR基因编码蛋白与其他植物上的该基因编码蛋白具有相同的保守结构域TIR,属于TIR-NB-LRR类型抗病基因家族,但不同植物间具有不同程度的同源性。SOPMA在线网站分析(图2b)显示,FtTIR蛋白二级结构包含41.36% α-螺旋、34.03%不规则卷曲、15.71%β-折叠及8.90% β-转角。三级结构预测(图2c)显示,FtTIR蛋白与模板c5ku7B的相似性达33%,与二级结构预测结果基本一致。利用Phyre2预测FtTIR的三级结构,显示FtTIR与TIR-NB-LRR型抗性蛋白rpv1(c5ku7B)的相似性达100%,与二级结构预测结果一致。系统发育分析(图2d)显示,FtTIR蛋白与藜麦XP_021726625.1和赤豆XP_047164412.1亲缘关系较近,属于同一分支。
图2

图2
FtTIR基因的生物信息学分析
(a) FtTIR与其他植物TIR蛋白的多重比对;(b) FtTIR蛋白二级结构预测:α-螺旋(蓝色)、延伸链(红色)、β-转角结构(绿色)、无规则卷曲(紫色);(c) FtTIR蛋白三级结构预测;(d) FtTIR蛋白系统进化树分析
Fig.2
Bioinformatics analysis of FtTIR gene
(a) Multiple alignment of FtTIR with other plant TIR proteins; (b) The secondary structure prediction of FtTIR protein:α-helix (blue), extended strand (red), β-turn (green), random coil (purple); (c) Thetertiary structure prediction of FtTIR protein; (d) Phylogenetic tree analysis of FtTIR protein
2.2 苦荞FtTIR基因的表达模式分析
qRT-PCR结果(图3)表明,FtTIR基因在立枯丝核菌侵染下总体表达上调,但苦荞根、茎和叶中FtTIR基因存在不同程度的诱导表达,且在根中的表达量普遍较低,在茎中的表达量较高。在茎中FtTIR基因的表达量在接菌后6h时达到峰值,与不接菌相比表达量上调约7倍。在根中FtTIR基因的表达量在接菌后48h时达到峰值,且表达量仅上调0.5倍左右。在叶中FtTIR基因的表达量在接菌后24h时达到峰值,表达量上调约9倍。
图3
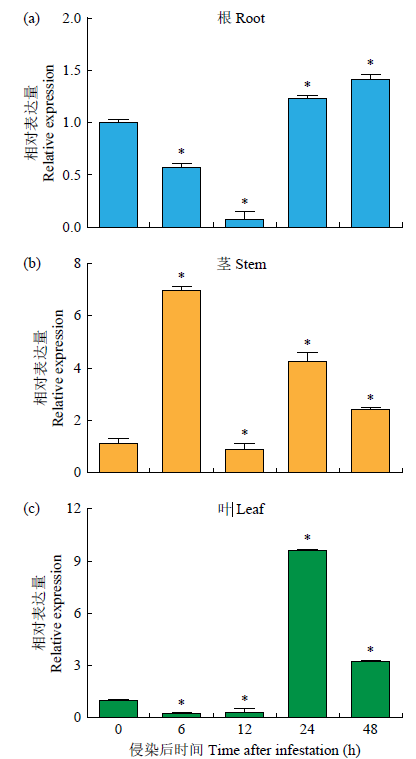
图3
立枯丝核菌侵染下FtTIR基因在苦荞不同器官中的表达模式
“*”表示在0.05水平下有显著性差异,下同
Fig.3
Expression patterns of FtTIR genes in different organs of tartary buckwheat under infestation with R.solani
“*”indicates significantly different at the 0.05 level, the same below
2.3 FtTIR过表达拟南芥的抗病性鉴定
2.3.1 过表达拟南芥叶片接种立枯丝核菌表型
选取生长状态相同的3周龄的转基因和野生型拟南芥叶片进行立枯丝核菌接种(图4),在正常条件下,野生型(WT)与过表达拟南芥株系(FtTIR-OE)之间无明显差异。而在立枯丝核菌侵染后,过表达拟南芥株系叶片的抗病性较WT强。
图4

图4
转基因拟南芥的抗病性表型验证
对照:不作处理;WT:野生型拟南芥;FtTIR-OE:FtTIR过表达拟南芥。下同
Fig.4
Phenotypic verification of disease resistance in transgenic Arabidopsis
Control: no treatment; WT: wild-type Arabidopsis; FtTIR-OE: overexpression of FtTIR in Arabidopsis. The same below
2.3.2 DAB染色试验
选取3周龄的FtTIR-OE和WT拟南芥叶片进行立枯丝核菌侵染,对照组不作侵染。经28℃培养箱培养24h后,用DAB染色液对离体叶片染色20min,再用叶绿体脱色液脱色至叶片透明。如图5所示,对照组FtTIR-OE和WT拟南芥叶片无明显差异,而被立枯丝核菌侵染的叶片出现大量的病斑,呈棕褐色。在立枯丝核菌侵染24h时,经DAB染色后,FtTIR-OE拟南芥株系叶片的病斑明显少于WT。这表明FtTIR基因对立枯丝核菌的侵害具有一定的抵抗作用。
图5

图5
转基因拟南芥的抗病性DAB染色验证
Fig.5
DAB staining for resistance of transgenic Arabidopsis
2.4 立枯丝核菌侵染FtTIR过表达拟南芥的表达模式
采用qRT-PCR方法检测FtTIR-OE和WT拟南芥在立枯丝核菌侵染下FtTIR基因的表达模式,以Atactin为内参。如图6a所示,FtTIR-OE转基因株系中FtTIR基因的表达量较WT显著提高,而在立枯丝核菌侵染后FtTIR-OE转基因株系(OE-RS)中FtTIR基因的表达量较不侵染时(OE)的表达量显著提高。
图6
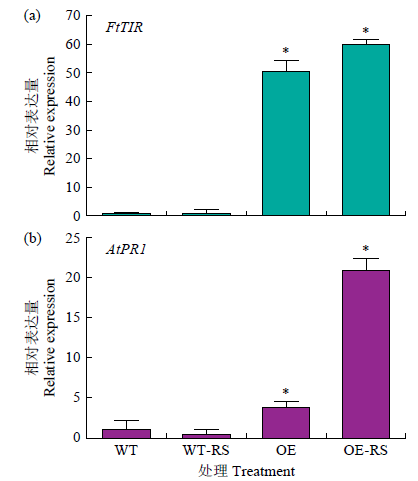
图6
转基因拟南芥中的表达模式分析
WT:野生型叶片未处理;WT-RS:野生型叶片受立枯丝核菌侵染;OE:FtTIR过表达拟南芥叶片未处理;OE-RS:FtTIR过表达拟南芥叶片受立枯丝核菌侵染。下同
Fig.6
Analysis of expression patterns in transgenic Arabidopsis
WT: Untreated wild type leaves; WT-RS: Wild-type leaves infected by R.solani; OE: Untreated FtTIR overexpressed Arabidopsis leaves; OE-RS: Overexpressed Arabidopsis leaves infected by R.solani. The same below
进一步研究FtTIR在苦荞立枯病抗病信号通路中的作用,通过qRT-PCR检测拟南芥叶片中病程相关基因AtPR1的表达水平,结果表明转基因拟南芥中AtPR1基因的表达量显著高于WT(图6b),同时在立枯丝核菌侵染下,AtPR1基因的表达量显著增加。
2.5 拟南芥抗病性相关生理生化指标分析
图7
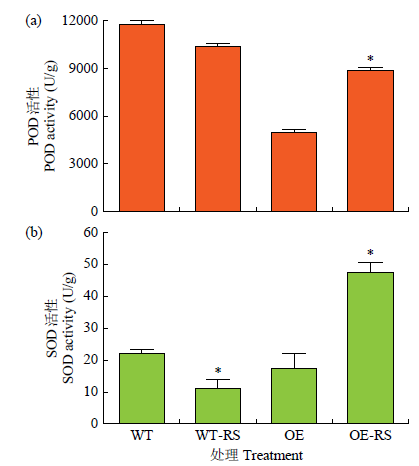
图7
转基因拟南芥生理生化指标分析
Fig.7
Analysis of biochemical indices in transgenic Arabidopsis
3 讨论
为了适应极端环境,植物进化出一系列防御机制来抵抗病原菌的侵害[15]。TIR抗病基因在植物的抗病信号通路中发挥着至关重要的作用[16]。荞麦的药用价值和经济价值日益显著,种植面积不断扩大,而其生产受立枯病影响日益严重[3,17]。立枯病是由立枯丝核菌感染造成的,用立枯丝核菌感染苦荞植株后,90%的植株出现坏死的褐色病变[18]。到目前为止,关于荞麦的立枯病抗病机制尚未明确。为解析苦荞对立枯丝核菌的抗病性,本研究从苦荞品种“品苦1号”中克隆出FtTIR基因,生物信息学分析显示其具有TIR家族特有的TIR结构域。在苦荞苗期,立枯丝核菌侵染后的qRT-PCR分析表明,FtTIR基因受立枯丝核菌胁迫的诱导;组织特异性分析表明,FtTIR基因在根、茎和叶中有不同的表达量,参与不同的发育阶段。
TIR基因的作用机制是通过与病原菌结合,将信号转递给转导因子[4]。转导因子与免疫反应基因的启动子相结合,诱导免疫反应基因表达,从而对病原菌产生抗性[18]。在烟草中,TIR基因的突变导致转基因烟草对烟草花叶病毒的抗性消失,且病毒的扩散速度大幅提升,说明TIR基因参与了烟草花叶病毒的信号转导[19]。在拟南芥中,TIR基因的突变导致转基因拟南芥在没有病原菌效应蛋白诱导的情况下也可以引起细胞坏死[11,20]。同样,在亚麻中TIR基因突变也导致转基因植株对条锈病的抗性消失[21]。这些研究证实,TIR基因与病原菌的效应分子识别有关,但TIR蛋白是否直接与病原菌效应蛋白结合还有待进一步研究证实。此外,TIR基因具有调节信号转导的功能[22]。对拟南芥中TIR基因的研究[20,23]发现,TIR基因具有负调节抗性信号传导的功能,因为TIR基因的突变导致转基因拟南芥的防御反应一直处于激活状态。同样,过表达马铃薯TIR基因导致植株的病菌防御反应降低[24]。与上述研究矛盾的是,有研究[25]发现,TIR基因具有正调节功能。将TIR基因的LRR结构域突变后,植株的抗性反应消失,说明LRR结构域具有正调控信号转化功能。因此,TIR是否作为植物抗病反应信号转导的负调控因子仍需进一步研究证实。
本研究中,在拟南芥中异源表达FtTIR后显著提高了拟南芥叶片对立枯丝核菌的抗病性。生理指标检测分析发现,其抗病性的提高与植物体内SOD和POD活性的增加有关。此外,在立枯丝核菌侵染过表达拟南芥中病程相关基因AtPR1的表达水平也显著提高,这种病程相关蛋白激活了植株的防御机制,从而使植物最大程度减少损伤。本研究结果表明,FtTIR在提高苦荞对立枯丝核菌抗病性中发挥着积极作用。本研究证实了TIR基因通过激活病程相关基因PR1的表达来提高植株的抗病性,是苦荞立枯病抗性的正向调节因子,该结果为后续深入研究TIR基因调控苦荞立枯病抗病分子机制奠定了基础。
4 结论
从苦荞中克隆出TIR-NB-LRR类型的抗病基因FtTIR,在立枯丝核菌侵染24h后,其在苦荞根、茎和叶中表达量均有显著增加,在茎中表达量最高,表明FtTIR基因受立枯丝核菌诱导表达。过表达拟南芥离体叶片侵染和DAB染色结果说明,FtTIR基因具有明显的抗病性。在立枯丝核菌侵染条件下,FtTIR转基因拟南芥植株的POD和SOD活性较WT显著提高,其抗病能力显著高于对照。
参考文献
植物抗病蛋白研究进展
DOI:10.11983/CBB17148
[本文引用: 1]

为了应对外界复杂的环境变化, 植物进化出一套复杂而精细的免疫应答调控机制。植物抗病蛋白能够特异地识别病原微生物分泌的效应蛋白, 触发免疫响应以对抗病原微生物的侵扰。该文综述了植物抗病蛋白的结构与功能及对病原菌的识别方式、在免疫响应过程中抗病蛋白的动态平衡机制及其介导的防御反应信号转导。开展植物抗病蛋白研究可为定向培育抗病作物奠定理论基础。
TNL genes in peach: insights into the post-LRR domain
HIGS-SsCCS转基因拟南芥的菌核病抗性鉴定
DOI:10.3864/j.issn.0578-1752.2020.04.008
[本文引用: 1]
【目的】菌核病是由核盘菌(Sclerotinia sclerotiorum)引起的一类真菌病害,核盘菌寄主范围广泛,严重危害多种作物的品质。本研究利用寄主诱导基因沉默(HIGS)的方法在寄主中诱导核盘菌致病相关基因的沉默从而增强寄主的菌核病抗性,为菌核病抗病育种提供新的思路。【方法】铜锌超氧化物歧化酶是一种重要的抗氧化剂,以核盘菌铜锌超氧化物歧化酶铜伴侣基因(copper chaperone for copper/zinc superoxide dismutase,SsCCS)为靶基因,通过生物信息学分析该基因的结构特点,并利用MEGA6.0软件构建系统发育树;通过分别比对拟南芥及核盘菌基因组,选择特异的干扰片段进行扩增;采用农杆菌介导的浸花序法,将HIGS载体转入拟南芥Col-0,通过DNA鉴定以及标记筛选出稳定的HIGS-CCS转基因拟南芥;选取4—5周龄的HIGS-CCS转基因拟南芥植株叶片接种核盘菌野生菌株1980,于接种24 h后统计病斑面积,分析转基因株系的菌核病抗性;通过qRT-PCR分析核盘菌侵染转基因植株过程中SsCCS的表达情况;同时在接种6、12、24 h后利用DAB染色的方法检测转基因植株与核盘菌互作过程中H<sub>2</sub>O<sub>2</sub>的积累。【结果】生物信息学分析结果表明,SsCCS(SS1G_00102)全长为1 010 bp,编码序列长759 bp,共编码253个氨基酸,该蛋白分子质量为27 176.96 Da,等电点(PI)为5.04,与灰霉病菌BcCCS(EDN25358)亲缘关系最近,氨基酸同源性达到87%,与拟南芥AtCCS(AT1G12520.1)亲缘关系较远;通过与核盘菌以及拟南芥基因组比对,选择314 bp特异干扰片段,成功构建SsCCS的HIGS表达载体,转化拟南芥。T<sub>1</sub>及T<sub>2</sub>代转基因拟南芥接种核盘菌24 h后的病斑面积均小于野生型拟南芥。从T<sub>2</sub>代中获得3个稳定表达的T<sub>3</sub>代HIGS-CCS转基因拟南芥株系:HIGS-CCS-5、HIGS-CCS-8、HIGS-CCS-13;与野生型拟南芥相比,转基因拟南芥在接种核盘菌24 h后,病斑面积显著减小46%—61%;qRT-PCR结果显示核盘菌在侵染转基因植株过程中,SsCCS的表达量显著低于侵染野生型拟南芥。DAB染色结果表明,在侵染过程中,转基因拟南芥植株中H<sub>2</sub>O<sub>2</sub>的积累明显少于野生型拟南芥,ROS水平降低。【结论】利用HIGS方法在拟南芥中干扰核盘菌SsCCS的表达能够显著提高拟南芥的抗病性,研究结果为油菜等农作物菌核病抗病改良提供了参考。
Comparative analysis of the flax immune receptors L6 and L 7 suggests an equilibrium-based switch activation model
DOI:10.1105/tpc.15.00303
PMID:26744216
[本文引用: 1]
NOD-like receptors (NLRs) are central components of the plant immune system. L6 is a Toll/interleukin-1 receptor (TIR) domain-containing NLR from flax (Linum usitatissimum) conferring immunity to the flax rust fungus. Comparison of L6 to the weaker allele L7 identified two polymorphic regions in the TIR and the nucleotide binding (NB) domains that regulate both effector ligand-dependent and -independent cell death signaling as well as nucleotide binding to the receptor. This suggests that a negative functional interaction between the TIR and NB domains holds L7 in an inactive/ADP-bound state more tightly than L6, hence decreasing its capacity to adopt the active/ATP-bound state and explaining its weaker activity in planta. L6 and L7 variants with a more stable ADP-bound state failed to bind to AvrL567 in yeast two-hybrid assays, while binding was detected to the signaling active variants. This contrasts with current models predicting that effectors bind to inactive receptors to trigger activation. Based on the correlation between nucleotide binding, effector interaction, and immune signaling properties of L6/L7 variants, we propose that NLRs exist in an equilibrium between ON and OFF states and that effector binding to the ON state stabilizes this conformation, thereby shifting the equilibrium toward the active form of the receptor to trigger defense signaling. © 2016 American Society of Plant Biologists. All rights reserved.
Multiple domain associations within the Arabidopsis immune receptor RPP 1 regulate the activation of programmed cell death
DOI:10.1371/journal.ppat.1005769 URL [本文引用: 1]
Expression of RPS4 in tobacco induces an AvrRps4-independent HR that requires EDS1, SGT1 and HSP90
DOI:10.1111/tpj.2004.40.issue-2 URL [本文引用: 2]
中国荞麦属植物分类学的修订
DOI:10.13430/j.cnki.jpgr.20181210001
[本文引用: 1]
荞麦是一种蓼科(Polygonaceae family)荞麦属(Fagopyrum Mill)的古老作物,在全世界,尤其是东亚及东欧地区广泛种植。数千年以来,荞麦都是中国凉山等高海拔地区的主要粮食作物,其营养丰富,其中富含的芦丁等生物活性成分还具有抗氧化、抗肿瘤的功效。荞麦的栽培种甜荞和苦荞已经被广泛传播,但野生荞麦却只在特定区域分布。中国西南地区,包括四川,云南,贵州各省以及西藏自治区,是公认的世界荞麦属植物的起源中心,分布中心和多样化中心,荞麦种质资源非常丰富。目前已有文献报导的荞麦属植物超过 20 种,他们中的绝大多数已在中国西南地区被发现,该地区的地理环境非常复杂,孕育了极具多样性的野生荞麦资源。但截至目前,这些已报导的种还没有经过科学的归类,同种异名的现象比较严重,本研究团队从 2004 年至今,在中国西南地区进行了多年多次的野外考察和标本采集工作,并在室内进行花和果实的解剖观察,获得了大量的荞麦属植物分类学数据。基于这些数据和荞麦染色体数量分析,经馆藏标本和模式照片的对比,并综合国内外相关文献,本文对中国荞麦属植物进行了分类学修订,将产自中国的荞麦属28 个种归并为21个:即将Fagopyrum megaspartaniumQ. F. Chen,F. pilus Q. F. Chen 并入 F. cymosum (Trrev)Meisn 作为异名;F. pleioramosum Ohnishi,F. wenchuanense J.R.Shao 并入F. caudatum (Samuelsson) A. J. Li 作为异名;F. callianthum Ohnishi 并入 F.qiangcai D.Q.Ba 作为异名;F. zuogongense Q.F.Chen 并入 F. homotropicum Ohnishi 作为异名;F. longzhoushanense J.R.Shao 并入 F. pugense T.Yu 作为异名。重新制定了 21 个种的分种检索表。本文将有助于重新认识荞麦属植物的进化地位,并为后续进一步探究属中各个种的亲缘关系奠定了基础。
荞麦轮纹病抗性鉴定方法的建立及荞麦抗病种质资源的筛选
DOI:10.11924/j.issn.1000-6850.casb16120120
[本文引用: 2]
为建立荞麦轮纹病的抗性鉴定方法和筛选可利用的抗轮纹病的荞麦种质资源,本研究以TP2 和KP14 为供试材料,在健康荞麦叶片正面针刺处理,然后用荞麦轮纹病的病原菌菌饼进行接种;利用建立的抗性鉴定方法,对50 份荞麦种质资源进行抗病性评价。结果显示:针刺5 针接种叶片病斑出现较快,且比针刺1 针、针刺3 针的叶片病斑大,针刺5 针的病情指数最高。50 份荞麦种质资源的病情指数有差异,其中YZ-18 的病情指数为29.63,为高抗种质资源,YZ-2、YZ-5、YZ-13 和YZ-9 的病情指数均低于40,为中抗种质资源,其余为感病种质资源。因此,接种前叶片正面针刺5 针的方法为最佳的抗性鉴定方法。50份抗性评价的荞麦种质资源中,筛选出5份抗病种质资源。
The Arabidopsis RPS4 bacterial-resistance gene is a member of the TIR-NBS-LRR family of disease-resistance genes
DOI:10.1046/j.1365-313X.1999.t01-1-00600.x URL [本文引用: 1]
Assessment of disease resistance and high yielding traits of common buckwheat genotypes in subtropical climate of Nepal
DOI:10.3329/sja.v18i1.48388
URL
[本文引用: 1]
Twelve common buckwheat (Fagopyrum esculentum Moench) genotypes received from Hill Crops Research Program, Kabre, Dolakha were evaluated for resistance against major foliar fungal diseases and high yielding traits at the research field of National Maize Research Program, Rampur, Chitwan during winter seasons of two consecutive years 2017 and 2018. The design of the experiment was Randomized Complete Block having three replications. The unit plot size was 3m x 2m with 25cm row to row spacing and continues plant to plant spacing was maintained and net harvested plot was 6 sq meters. The experiment was planted at last week of October in both years. The recommended dose of fertilizer was 30:30:0 for N:P:K kg ha-1 respectively and seed rate 50 kg ha-1. Disease data were recorded for downy mildew (Perenospora fagopyri), powdery mildew (Erysiphepolygoni) and botrytis leaf blight (Botrytis cinerea) severity on 20 randomly tagged plants/plot. The yield and yield attributing traits were recorded. Buckwheat genotypes differed significantly (p<0.05) on disease severity, grain yield and yield attributing traits. Genotypes CBBP-01, KIF-72-22-520, GF5283, ACC#2234 and ACC#2213 were resistant to powdery mildew, downy mildew and botrytis leaf blight and also resulted in higher grain yield. Grain yield had found strong negative correlation with the fungal disease severity of buckwheat. These genotypes might be useful for the development of major foliar fungal disease resistant high yielding common buckwheat variety in inner terai region of Nepal.
\nSAARC J. Agri., 18(1): 143-152 (2020)
立枯丝核菌毒素的研究进展
DOI:10.11869/j.issn.100-8551.2020.10.2219
[本文引用: 2]
立枯丝核菌(Rhizoctonia solani kühn)寄主范围广且腐生性强,主要危害禾本科、茄科、豆科和十字花科等43科260多种植物,造成水稻、玉米和马铃薯等作物产量严重减少以及品质下降。目前种质库中尚未发现高水平的抗源,因此无法借助转基因手段培育抗立枯丝核菌的新材料。立枯丝核菌致病因子主要为毒素和细胞壁降解酶,基于病原菌和植物互作关系,本文着重阐述了立枯丝核菌毒素提取工艺的发展、成分鉴定、生物活性及致病机理,旨在为进一步深入研究立枯丝核菌及其相关病害的有效防治提供一定的理论依据。
The Arabidopsis thaliana TIR-NB-LRR R-protein, RPP1A; protein localization and constitutive activation of defence by truncated alleles in tobacco and Arabidopsis
DOI:10.1111/tpj.2006.47.issue-6 URL [本文引用: 2]
Structural and functional analysis of a plant resistance protein TIR domain reveals interfaces for self-association, signaling, and autoregulation
Mutational analysis of the Arabidopsis nucleotide binding site: leucine-rich repeat resistance gene RPS2
Resistance in transgenic potato expressing the potato leafroll virus coat protein gene
DOI:10.1094/MPMI-3-301 URL [本文引用: 1]
Evidence for a role of the N terminus and leucine-rich repeat region of the Mi gene product in regulation of localized cell death
DOI:10.1105/tpc.12.8.1319 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}