


三酰甘油(triacylglycerol,TAG)是植物油脂的主要贮存形式,作为能量来源在种子萌发、叶片代谢和有性生殖等生理过程中发挥重要作用[1-2]。二酰甘油酰基转移酶(diacylglycerol acyltransferas,DGAT)广泛参与TAG合成过程,是催化TAG合成的关键酶和唯一限速酶[3-4]。根据结构差异和底物特异性,DGAT分为4种亚型,分别为DGAT1、DGAT2、DGAT3和WSD/DGAT。DGAT1和DGAT2作为其主要亚型,广泛分布于真核生物中。DGAT1和DGAT2具有相对独立的进化轨迹,功能趋于同质化[5]。花生基因AhDGAT1.1在野生型烟草中过量表达可增加种子脂肪酸含量[6]。Zhou等[7]通过构建瞬时表达系统发现,平行表达的拟南芥AtDGAT1和AtDGAT2基因均可增加烟草叶片中TAG含量,但AtDGAT2相比AtDGAT1拥有更高的转化效率。DGAT3是一种可溶性酶,可作为TAG合成备选途径发挥作用[8]。相关研究[9]表明,DGAT3被描述为DGAT金属蛋白,其蛋白功能与铁的可获得性有关。陶芬芳[10]发现,甘蓝型油菜基因BnDGAT3能够改变拟南芥和酵母的脂肪酸成分,且具有底物偏向性。WSD/DGAT是新发现的DGAT亚型,作为双功能酶催化蜡酯及TAG合成,但DGAT活性较低[11]。DGAT基因不仅对植物油脂合成具有重要作用,还广泛参与植株建成和胁迫响应等过程[12-13]。Lu等[14]研究表明,拟南芥DGAT突变体与野生型相比具有显著的胁迫敏感性,同时其种子萌发和幼苗生长等均受到显著影响。Siloto等[15]研究也证实,DGAT基因的表达抑制会导致种子发芽率降低,并伴有畸形植株产生。
玉米、水稻、小麦、高粱和谷子是我国禾谷类主要粮食作物,主要用于粮饲供给和工业加工。研究[16]表明,高油品种相比普通品种拥有更高的营养价值和饲料转化率。此外,由于化学结构上的相似性,植物油可作为石油的可再生替代品之一[17]。因此,培育高油品种具有巨大的经济潜力和广泛的社会意义。基因组测序工作的逐步完成,为功能基因组学构建了数据平台[18]。关于DGAT基因家族的研究,多集中于大豆和花生等油料作物,对禾谷类粮食作物鲜有报道。为明确禾谷类粮食作物DGAT基因家族的表达特征,本研究利用生物信息学方法对上述5种禾谷类作物DGAT基因家族进行全基因组鉴定,分析其亚型差异和功能特点,为DGAT成员生物学功能研究奠定理论基础。
1 材料与方法
1.1 试验材料
供试玉米(Zea mays L.)品种为自交系黄早4,利用15% NaClO对种子消毒7min,蒸馏水洗净,随后进行黑暗条件下的萌发处理。发芽培养12h后移栽至光照培养箱,在Hoagland培养液中进行水培,培养箱条件设置为16h光照/8h黑暗。待植株长至3叶1心期时,进行低温(4℃)、盐(200mmol/L NaCl)、脱落酸(ABA,100μmol/L)和干旱(20% PEG 8000)胁迫处理,于不同处理时间(0、6、12、24h)取根部,液氮速冻备用[13]。
1.2 序列获取及特征分析
利用已知花生DGAT成员序列(GI:XP_ 016202335.1、GI:015950782.1、GI:AAX62735.1、GI:XP_015964946.1)[19]在玉米(Zea mays L.)、水稻(Oryza sativa L.)、小麦(Triticum aestivum L.)、高粱(Sorghum bicolor L. Moench)和谷子(Setaria italica)基因组中进行BLASTP搜索,利用SMART(
1.3 进化树构建及基序、结构域分析
通过MEME网站(
1.4 蛋白互作网络构建
利用STRING数据库(
1.5 顺式作用元件预测及基因结构分析
DGAT成员基因序列和编码序列来自Ensembl Plants数据库(
1.6 DGAT基因表达模式分析
利用NCBI数据库(
1.7 DGAT基因qRT-PCR分析
使用Trizol公司试剂盒提取玉米根部在遭遇低温、干旱、盐和ABA胁迫时不同时期的RNA,使用PrimeScript RT Reagent Kit With gDNA Eraser将RNA反转录为cDNA,内参基因选择ZmActin,PCR反应体系为95℃ 60s;95℃ 15s,55℃ 30s,72℃ 60s,共40个循环。每个样品重复3次,通过2‒ΔΔCT法处理数据,使用TBtools进行热图绘制。
2 结果与分析
2.1 DGAT序列鉴定及特征分析
利用已知DGAT成员序列,对玉米、水稻、小麦、高粱和谷子进行全基因组扫描,E值过滤后通过SMART进行结构域鉴定,最终确定DGAT成员,其中玉米7个,水稻7个,小麦26个,高粱9个,谷子10个。由表1可知,DGAT成员氨基酸长度为140~565,分子量为15.71~62.37kDa,等电点为6.30~10.57,不稳定系数为32.44~54.99,脂肪系数为78.96~106.90,亲水性总均值为-0.251~ 0.500。二级结构预测结果表明,DGAT蛋白以α-螺旋为主,无规则卷曲次之,β-转角最少。经比对发现,DGAT蛋白理化性质在亚型间并未呈现出明显差异,主要为高脂肪疏水蛋白,具有弱碱性和不稳定性特点。亚细胞定位结果显示,DGAT1亚型成员均定位到质膜中;DGAT2成员中有70%定位到质膜,20%定位到叶绿体,10%定位到细胞质;WS/DGAT(即OsWSD-1和OsWSD-2等)成员中有44%定位到质膜,20%定位到细胞质,20%定位到叶绿体,16%定位到内质网。不同定位结果表明,DGAT成员参与不同生物学过程。
表1 DGAT家族成员信息
Table 1
| 物种 Species | 基因名 Genetic name | 染色体位置 Chromosome position | 氨基酸 长度 Amino acid length | 分子量 Molecular weight (kDa) | 理论 等电点 Isoelectric point | 不稳定系数 Coefficient of instability | 脂肪系数 Fat coefficient | 亲水性 总均值 Hydrophilic total mean | α-螺旋 α- spiral (%) | 延伸链 Extended chain (%) | β-转角 β- corner (%) | 无规则 卷曲 Random coil (%) | 亚细胞定位 Subcellular localization |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 水稻 Oryza sativa | OsDGAT1-1 | chr5.5971951-5978628 | 538 | 59.60 | 9.15 | 48.49 | 99.01 | 0.265 | 48.14 | 10.22 | 3.72 | 37.92 | 质膜 |
| OsDGAT1-2 | chr6.21677541-21684368 | 477 | 55.11 | 9.43 | 44.61 | 106.90 | 0.309 | 50.31 | 13.00 | 4.40 | 32.29 | 质膜 | |
| OsDGAT2-1 | chr2.29599167-29601738 | 338 | 37.55 | 9.71 | 35.13 | 95.18 | 0.299 | 36.98 | 21.01 | 9.17 | 32.84 | 质膜 | |
| OsDGAT2-2 | chr6.12809778-12812419 | 340 | 37.77 | 9.34 | 34.22 | 98.94 | 0.311 | 38.24 | 19.12 | 7.35 | 35.29 | 质膜 | |
| OsWSD-1 | chr1.28033034-28051826 | 484 | 52.75 | 7.27 | 35.77 | 85.91 | 0.007 | 36.78 | 16.74 | 4.54 | 41.94 | 细胞质 | |
| OsWSD-2 | chr1.32484736-32489971 | 531 | 58.21 | 6.77 | 45.88 | 87.87 | -0.144 | 38.42 | 17.89 | 3.77 | 39.92 | 质膜 | |
| OsWSD-3 | chr5.27660691-27664963 | 565 | 62.37 | 9.11 | 53.17 | 86.88 | -0.191 | 38.94 | 14.87 | 3.18 | 43.01 | 内质网 | |
| 谷子 Setaria italica | SiDGAT1-1 | chr3.7546566-7553318 | 521 | 58.43 | 9.13 | 48.63 | 102.42 | 0.311 | 47.60 | 10.75 | 4.41 | 37.24 | 质膜 |
| SiDGAT1-2 | chr4.36212704-36219516 | 495 | 56.79 | 9.51 | 48.61 | 104.38 | 0.298 | 46.26 | 11.52 | 3.64 | 38.59 | 质膜 | |
| SiDGAT2-1 | chr1.36275566-36279052 | 273 | 30.03 | 9.91 | 41.83 | 101.83 | 0.469 | 33.70 | 19.78 | 7.69 | 38.83 | 质膜 | |
| SiDGAT2-2 | chr4.16556591-16559685 | 364 | 40.31 | 10.66 | 54.99 | 85.49 | -0.077 | 29.12 | 18.41 | 7.69 | 44.78 | 质膜 | |
| SiWSD-1 | chr2.1723994-1726391 | 477 | 52.20 | 8.57 | 43.36 | 98.99 | 0.095 | 40.25 | 17.61 | 3.14 | 38.99 | 质膜 | |
| SiWSD-2 | chr3.11005742-11010410 | 273 | 30.04 | 9.91 | 41.83 | 101.83 | 0.469 | 33.70 | 19.78 | 7.69 | 38.83 | 内质网 | |
| SiWSD-3 | chr5.33312705-33316538 | 538 | 57.92 | 7.67 | 44.54 | 87.10 | 0.013 | 41.64 | 13.57 | 3.72 | 41.08 | 细胞质 | |
| SiWSD-4 | chr5.37959858-37963149 | 491 | 54.05 | 9.09 | 39.99 | 94.38 | -0.034 | 40.94 | 15.07 | 3.46 | 40.53 | 叶绿体 | |
| SiWSD-5 | chr8.35767324-35770211 | 439 | 47.88 | 8.77 | 43.46 | 99.00 | 0.080 | 36.90 | 15.72 | 3.64 | 43.74 | 质膜 | |
| SiWSD-6 | chr9.45940206-45942823 | 515 | 56.14 | 6.68 | 40.82 | 93.44 | 0.000 | 38.45 | 16.31 | 3.69 | 41.55 | 内质网 | |
| 小麦 Triticum aestivum | TaDGAT1-1 | chr1A.148953747-148959478 | 513 | 57.74 | 8.92 | 51.01 | 99.61 | 0.295 | 48.34 | 11.70 | 4.29 | 35.67 | 质膜 |
| TaDGAT1-2 | chr1B.195590716-195596372 | 513 | 57.75 | 8.99 | 50.36 | 99.81 | 0.299 | 47.56 | 12.87 | 4.29 | 35.28 | 质膜 | |
| TaDGAT1-3 | chr1D.141705469-141711495 | 513 | 57.78 | 9.07 | 49.88 | 99.61 | 0.290 | 48.34 | 11.89 | 4.48 | 35.28 | 质膜 | |
| TaDGAT1-4 | chr7A.532701958-532710211 | 470 | 54.07 | 9.20 | 43.38 | 105.38 | 0.291 | 47.87 | 11.28 | 2.98 | 37.87 | 质膜 | |
| TaDGAT1-5 | chr7B.491164040-491172543 | 471 | 54.14 | 9.20 | 43.22 | 105.37 | 0.293 | 46.28 | 11.68 | 2.97 | 39.07 | 质膜 | |
| TaDGAT1-6 | chr7D.467917764-467926136 | 470 | 54.07 | 9.25 | 43.56 | 106.43 | 0.301 | 47.45 | 12.13 | 3.83 | 36.60 | 质膜 | |
| TaDGAT2-1 | chr1A.309910012-309913118 | 247 | 28.36 | 9.85 | 41.85 | 100.97 | 0.199 | 28.34 | 27.13 | 7.69 | 36.84 | 细胞质 | |
| TaDGAT2-2 | chr6A.506235347-506239101 | 455 | 50.25 | 10.66 | 40.57 | 82.79 | -0.208 | 29.01 | 18.02 | 5.27 | 47.69 | 叶绿体 | |
| TaDGAT2-3 | chr6B.550077872-550080884 | 472 | 52.14 | 10.57 | 47.88 | 78.96 | -0.251 | 28.60 | 15.25 | 8.05 | 48.09 | 质膜 | |
| TaDGAT2-4 | chr6D.365278335-365281853 | 325 | 36.50 | 9.79 | 33.08 | 103.17 | 0.273 | 37.85 | 20.62 | 5.85 | 35.69 | 质膜 | |
| TaDGAT2-5 | chr7A.14172212-14175945 | 334 | 37.25 | 9.08 | 43.90 | 90.78 | 0.275 | 34.14 | 19.76 | 6.29 | 39.82 | 质膜 | |
| TaDGAT2-6 | chr7A.195308047-195310553 | 336 | 37.36 | 9.05 | 40.72 | 99.29 | 0.226 | 33.93 | 20.54 | 9.23 | 36.31 | 细胞质 | |
| TaDGAT2-7 | chr7B.332923582-332926431 | 344 | 38.04 | 8.76 | 40.06 | 101.54 | 0.255 | 37.50 | 17.73 | 7.56 | 37.21 | 质膜 | |
| TaDGAT2-8 | chr7D.14836474-14841174 | 324 | 36.23 | 9.51 | 46.31 | 89.66 | 0.241 | 37.96 | 19.14 | 7.10 | 35.80 | 质膜 | |
| TaDGAT2-9 | chr7D.188821967-188824699 | 347 | 38.47 | 8.90 | 40.64 | 101.47 | 0.248 | 30.84 | 19.88 | 7.78 | 41.50 | 质膜 | |
| TaWSD-1 | chr2A.24154221-24157435 | 218 | 25.13 | 6.30 | 48.21 | 91.19 | -0.128 | 47.71 | 12.84 | 5.50 | 33.94 | 细胞质 | |
| TaWSD-2 | chr2D.1912608-1922943 | 496 | 55.43 | 9.67 | 44.73 | 96.67 | -0.027 | 40.73 | 16.13 | 2.42 | 40.73 | 叶绿体 | |
| TaWSD-3 | chr3A.9843405-9846523 | 495 | 55.06 | 8.98 | 46.92 | 95.29 | 0.032 | 39.19 | 16.77 | 3.03 | 41.01 | 质膜 | |
| TaWSD-4 | chr3A.11023281-11028199 | 501 | 56.02 | 9.04 | 52.97 | 94.31 | 0.001 | 37.92 | 15.97 | 2.59 | 43.51 | 质膜 | |
| TaWSD-5 | chr3B.5230206-5234785 | 334 | 37.25 | 9.08 | 43.90 | 90.78 | 0.275 | 34.13 | 19.76 | 6.29 | 39.82 | 质膜 | |
| TaWSD-6 | chr3B.6670118-6673010 | 499 | 55.56 | 8.98 | 45.90 | 93.55 | 0.003 | 38.28 | 17.03 | 2.81 | 41.88 | 叶绿体 | |
| TaWSD-7 | chr3B.13744964-13749320 | 504 | 56.45 | 8.87 | 52.07 | 93.00 | 0.016 | 37.50 | 17.26 | 2.58 | 42.66 | 质膜 | |
| TaWSD-8 | chr3B.481649743-481652877 | 509 | 56.58 | 8.76 | 40.81 | 98.45 | 0.019 | 41.45 | 16.50 | 4.13 | 37.92 | 质膜 | |
| TaWSD-9 | chr3D.4179986-4182963 | 496 | 55.35 | 9.08 | 44.33 | 92.56 | -0.011 | 38.91 | 16.13 | 3.02 | 41.94 | 叶绿体 | |
| TaWSD-10 | chr3D.370123379-370126365 | 432 | 48.08 | 6.52 | 38.86 | 95.25 | -0.007 | 44.21 | 15.51 | 3.94 | 36.34 | 细胞质 | |
| TaWSD-11 | chr6B.550077872-550080884 | 500 | 55.57 | 9.10 | 47.36 | 93.40 | 0.041 | 39.40 | 19.80 | 2.80 | 38.00 | 质膜 | |
| 玉米 Zea mays | ZmDGAT1-1 | chr6.108212462-108219350 | 197 | 23.65 | 9.32 | 37.25 | 101.37 | 0.471 | 60.41 | 11.68 | 3.55 | 24.37 | 质膜 |
| ZmDGAT1-2 | chr6.136606701-136613008 | 536 | 60.17 | 9.21 | 48.34 | 101.19 | 0.352 | 50.00 | 11.38 | 4.48 | 34.14 | 质膜 | |
| ZmDGAT2-1 | chr4.12951363-12952979 | 140 | 15.71 | 6.80 | 49.60 | 106.50 | 0.500 | 31.43 | 25.00 | 7.14 | 36.43 | 叶绿体 | |
| ZmDGAT2-2 | chr4.165057547-165060492 | 409 | 45.26 | 10.75 | 45.52 | 85.40 | -0.072 | 30.56 | 15.65 | 6.60 | 47.19 | 质膜 | |
| ZmDGAT2-3 | chr5.50715540-50717216 | 204 | 22.88 | 8.70 | 39.00 | 96.03 | 0.009 | 31.37 | 24.51 | 10.78 | 33.33 | 叶绿体 | |
| ZmDGAT2-4 | chr9.47487176-47490826 | 332 | 37.16 | 9.60 | 32.44 | 97.80 | 0.247 | 37.35 | 19.88 | 6.33 | 36.45 | 质膜 | |
| ZmWSD-1 | chr6.167699425-167706622 | 556 | 61.29 | 9.23 | 47.05 | 83.63 | -0.170 | 39.57 | 15.29 | 2.70 | 42.45 | 质膜 | |
| 高粱 Sorghum bicolor | SbDGAT1-1 | chr9.8916935-8923148 | 515 | 57.86 | 9.13 | 46.93 | 99.86 | 0.295 | 48.93 | 10.68 | 3.50 | 36.89 | 质膜 |
| SbDGAT1-2 | chr10.50099836-50118712 | 484 | 55.62 | 9.23 | 44.56 | 102.75 | 0.294 | 51.24 | 11.57 | 2.48 | 34.71 | 质膜 | |
| SbDGAT2-1 | chr4.60698402-60701452 | 334 | 37.08 | 9.81 | 37.91 | 98.95 | 0.320 | 35.33 | 18.56 | 7.19 | 38.92 | 质膜 | |
| SbDGAT2-2 | chr8.4821575-4825729 | 290 | 33.34 | 9.36 | 44.87 | 81.97 | 0.169 | 33.79 | 24.83 | 7.59 | 33.79 | 叶绿体 | |
| SbDGAT2-3 | chr10.19611856-19615565 | 332 | 37.50 | 9.76 | 39.99 | 91.63 | 0.158 | 37.35 | 19.58 | 8.13 | 34.94 | 质膜 | |
| SbWSD-1 | chr2.1839607-1843547 | 494 | 54.62 | 9.11 | 52.05 | 93.99 | -0.052 | 37.65 | 17.41 | 3.85 | 41.09 | 质膜 | |
| SbWSD-2 | chr3.63801546-63804967 | 372 | 41.50 | 9.00 | 40.16 | 95.94 | 0.048 | 41.40 | 17.20 | 3.49 | 37.90 | 内质网 | |
| SbWSD-3 | chr6.9222487-9226442 | 468 | 51.05 | 7.20 | 37.29 | 97.14 | 0.060 | 34.19 | 17.31 | 2.78 | 45.73 | 细胞质 | |
| SbWSD-4 | chr9.56810016-56817272 | 554 | 61.07 | 9.34 | 45.02 | 81.44 | -0.175 | 39.89 | 15.34 | 3.25 | 41.52 | 质膜 |
2.2 DGAT进化树构建
为解读DGAT成员进化关系,以蒺藜苜蓿和大豆作为外类群进行进化树构建(图1)。结果表明,同一亚型DGAT成员在进化树中聚类为同一亚组,蒺藜苜蓿和大豆DGAT成员在此基础上单独聚类为一分支。Ⅰ-1、Ⅰ-2、Ⅱ-1和Ⅱ-2成员数量分别占进化树的23.73%、33.90%、3.39%和38.98%。根据拓扑结构,DGAT同源基因共24对,其中直系同源基因共12对,发生在高粱与谷子(2对)、高粱与玉米(5对)、水稻与谷子(1对)、大豆与蒺藜苜蓿(4对)之间;旁系同源基因共12对,其中玉米基因组中有1对,谷子基因组中有1对,小麦基因组中有10对。
图1

2.3 DGAT序列比对
为进一步解析DGAT各亚型之间差异,利用DNAMAN进行序列比对(图2a,仅展示部分代表成员)。结果发现,同亚型DGAT成员之间具有较高同源性,不同亚型成员之间同源性较低。其中,WS/DGAT亚型成员具有HHXXXDG活性位点,该位点被证明在蜡脂合成过程中发挥重要作用,同时参与TAG合成催化过程。DGAT2亚型成员包含PH模块、PR模块和GGE模块。其中,PH Block与酶活性有关,PR模块和GGE模块与底物识别、催化及低聚物形成有关[21-22]。DGAT3亚型成员包含2个DGAT3结构域和脂肪酸结合信号结构域和催化活性位点。DGAT1亚型成员包含酰基辅酶A结合区、催化活性位点SRLIIEN、SnRK1靶向位点、脂肪酸结合蛋白信号区、DAG结合区和识别内质网的信号区。部分区间位点保守性分析结果(图2b)表明,各亚型成员区间氨基酸位点具有较高的一致性,说明不同DGAT亚型成员可能存在分化。
图2

图2
DGAT氨基酸序列分析
a:DGAT序列比对,b:DGAT氨基酸位点分析
Fig.2
Analysis of DGAT amino acid sequence
a: DGAT sequence alignment, b: analysis of DGAT amino acid sites
2.4 DGAT基序和结构域分析
为解读DGAT亚型成员的进化特点,利用GSDS 2.0分析其基因结构,利用MEME分析其基序组成,采用TBtools进行整合。基因结构分析结果(图3a)显示,同亚型成员具有相似的剪切方式及外显子数量,但不同亚型成员间差异较大。蛋白基序分析结果显示,不同亚型成员具有特异的基序组成,不同亚型间不存在相同基序。DGAT3亚型成员并未检索到基序组成,说明DGAT3成员与其他亚型成员同源性较低。CDS基序分析结果显示,不同亚型成员具有特异的基序排列方式,其中motif 7、motif 12和motif 15几乎存在于所有亚型成员间。但DGAT3成员(MtDGAT3-1)同样没有检索到基序组成。为解读DGAT3成员差异,对2个DGAT3成员进行重新扫描(图3b),发现DGAT3成员具有保守的基序组成,但存在部分变异情况。结构域预测结果显示(图3c,研究物种中并未扫描到DGAT3成员,故仅展示其他亚型结构域组成),DGAT1成员均含有MBOAT结构域,DGAT2成员含有DAGAT结构域,WS/DGAT成员含有WES_ acyltransf和DUF1298结构域。其中,MBOAT被注释为膜结合O-酰基转移酶结构域,DAGAT注释为二酰甘油酰基转移酶结构域,WES_acyltransf被注释为蜡酯合酶样酰基辅酶A结构域,DUF1298为WS/DGAT特异的C端结构域。图3d展示了DGAT 3种亚型成员蛋白结构差异。
图3

图3
DGAT基因结构、蛋白结构、基序、结构域分析
(a) DGAT基因结构及基序分析,(b) DGAT3基序分析,(c) DGAT结构域分析,(d) DGAT三级结构
Fig.3
Genetic structure, protein sequence, gene sequence, domain of DGAT gene
(a) DGAT gene structure and motif analysis, (b) motif analysis of DGAT3, (c) DGAT domain analysis, (d) 3D structure of DGAT
2.5 DGAT蛋白功能注释及蛋白互作网络构建
为解读DGAT蛋白功能,以玉米DGAT成员为例构建了DGAT蛋白互作网络(图4)。发现该网络以ZmDGAT1-1和ZmDGAT1-2为核心节点构成,并包含糖基转移酶MGDG、非特异性脂质酰基水解酶Patatin和脂质代谢酶LCAT。但ZmWSD-1并未参与到该互作网络中,可能WS/DGAT蛋白拥有其独特的调节网络。
图4

图4
ZmDGAT蛋白互作网络
淡蓝色线段表示来自数据库,紫色表示试验证明,黄色表示文本挖掘,黑色表示共表达
Fig.4
ZmDGAT protein interaction network
The light blue line represents data from the database, the purple line represents experimental proof, the yellow line represents text mining, and the black line represents co-expression
2.6 DGAT启动子解析
为解读DGAT表达调控方式,利用TBtools软件绘制了顺式作用元件数量热图。结果(图5)显示,DGAT成员存在胁迫及植株建成等多种类型作用元件。其中,ABA响应元件、厌氧诱导元件、生长素响应元件、光响应元件、低温响应元件、茉莉酸响应元件大量分布在DGAT成员中,暗示DGAT成员广泛参与非生物胁迫过程。同时,胚乳特异性表达元件、种子特异性调控元件和细胞周期调控元件存在小部分成员中,暗示该部分成员具有相应的特殊功能。
图5
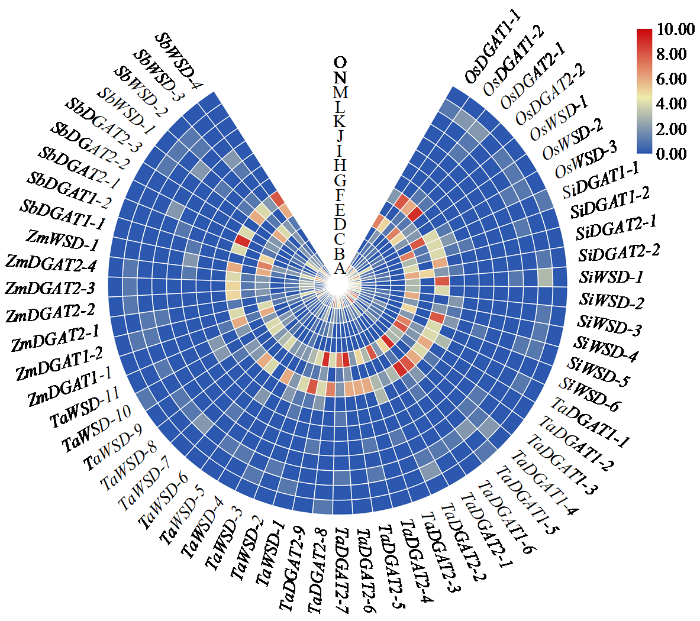
图5
DGAT启动子分析
A:ABA响应元件,B:厌氧诱导元件,C:生长素响应元件,D:防御与应激响应元件,E:光响应元件,F:低温响应元件,G:茉莉酸响应元件,H:分生组织表达,I:昼夜节律调控元件,J:赤霉素响应元件,K:胚乳特异性表达元件,L:种子特异性调控元件,M:玉米醇溶蛋白代谢调控元件,N:水杨酸响应元件,O:细胞周期调控元件
Fig.5
DGAT promoter analysis
A: ABA response element, B: anaerobic induction element, C: auxin response element, D: defense and stress response element, E: light response element, F: low temperature response element, G: jasmonic acid response element, H: meristem expression, I: circadian rhythm control element, J: gibberellin response element, K: endosperm specific expression element, L: seed specific control element, M: zein metabolism control element, N: salicylic acid response element, O: cell cycle control element
2.7 DGAT表达模式解析
为解读DGAT表达模式,利用已公布转录组数据构建DGAT成员在植株建成及逆境胁迫下的表达热图。图6a表明(TaWSD-1无匹配结果),大部分DGAT成员在根中高表达,在叶中表达量较低。同时存在部分基因(如TaWSD-5和TaWSD-11)在各组织或器官中均不表达,判断其可能具有特殊表达模式。由图6b可知,SbDGATs成员在植株遭遇ABA、干旱及弱碱胁迫时差异表达,在不同组织中具有特异性。如在遭遇弱碱胁迫处理时SbWSD-1在茎中特异表达,但在其他情况下不表达或表达量较低。SiDGATs同样存在此类现象,如SiWSD-6在正常光诱导时不表达,在遭遇其他胁迫时差异表达(图6c)。图6d表明,大部分TaDGATs在遭遇干旱胁迫时呈现先上升再下降的趋势。在遭遇高温及复合胁迫时则呈现持续下降趋势。同时存在部分基因具有特异表达模式,如TaWSD-10遭遇干旱胁迫下调表达,遭遇高温和复合胁迫上调表达,随着处理时间的上升,表达趋势与其他成员也具有明显差异。OsDGATs大部分基因在遭遇高温胁迫后上调表达,在遭遇低温时下调表达(图6e),说明DGAT成员在不同物种中具有不同表现。由图6f可知,ZmDGATs在不同组织中或遭遇不同程度胁迫处理时,同样存在特异表达现象。当植株遭遇淹没处理时,耐受品种(L3、L4)相较敏感品种(L1、L2)DGAT表达量较低。但存在特殊基因ZmWSD-1在L3中高表达,说明DGAT成员在不同品种间表达情况同样具有特异性。
图6
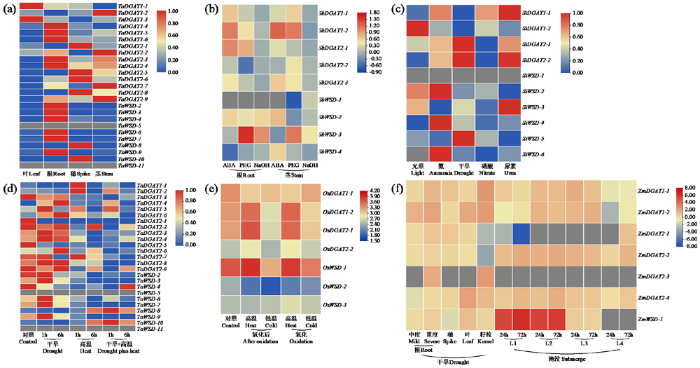
图6
DGAT基因家族表达模式分析
部分基因未匹配到数据,灰色代表基因不表达。(a):TaDGATs植株建成表达热图;(b):ABA、干旱、弱碱胁迫下SbDGATs在根和茎中表达热图;(c):氨、干旱、硝酸、尿素胁迫下SiDGATs表达热图;(d):高温、干旱、复合胁迫下TaDGATs表达热图;(e):不同氧化处理的高温、低温胁迫下OsDGATs表达热图;(f):干旱、淹没胁迫下ZmDGATs基因表达热图(L1-玉米品种B73,L2-玉米品种B97,L3-玉米耐受品种Mo18w,L4-玉米耐受品种M162w)
Fig.6
Analysis of the expression pattern of DGAT gene famil
Some genes were not matched to the data, and the gray color indicates that the genes are not expressed. (a): Heat map of TaDGATs plant built expression; (b): Heat map of SbDGATs expression in roots and stems under ABA, drought, weak alkaline stress; (c): Heat map of SiDGATs expression under light, ammonia, drought, nitrate, urea stress; (d): Heat map of TaDGATs expression under high temperature, drought, combined stress; (e): Heat and low temperature stress with different oxidative treatments OsDGATs expression heat map; (f): ZmDGATs gene expression heat map under drought and submergence stress (L1-maize variety B73, L2-maize variety B97, L3-maize tolerant variety Mo18w, L4-maize tolerant variety M162w)
2.8 DGAT基因qRT-PCR表达特征分析
ZmDGATs广泛响应非生物胁迫,转录水平表现不同特征。由图7可知,低温胁迫下,ZmDGAT1-1和ZmDGAT2-4下调表达,ZmWSD-1显著上调表达;盐胁迫下,ZmDGATs均上调表达,ZmDGAT1-2、ZmDGAT2-2和ZmWSD-1上调明显;ABA胁迫下,ZmWSD-1呈现先上升再下降的趋势,12h达到峰值,其他基因也具有较高表达量;干旱胁迫下,ZmDGAT1-1、ZmDGAT1-2、ZmDGAT2-2和ZmWSD-1显著上调表达,ZmDGAT2-4表达量变化不明显。
图7
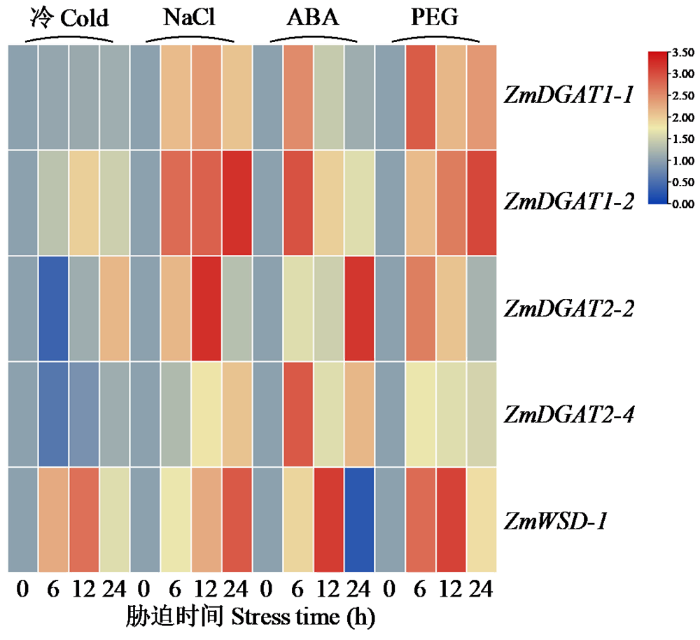
图7
ZmDGATs在非生物胁迫下qRT-PCR的表达分析
Fig.7
qRT-PCR expression analysis of ZmDGATs under abiotic stress
3 讨论
DGAT是植物三酰甘油合成途径的关键酶,对植物油脂的合成与储存具有重要意义。本研究通过对5种主要禾谷类作物DGAT成员鉴定分析,发现DGAT成员受多倍化影响,产生了一定程度的增减。小麦基因组中鉴定到26个DGAT家族成员,相较玉米(7个)、水稻(7个)、高粱(9个)和谷子(10个)具有明显差异,说明DGAT在进化过程中出现了全基因组复制现象,小麦作为六倍体植物所具有的高同源保留率可能是这种现象产生的原因之一[23]。进化树拓扑结构显示,DGAT同一亚型成员聚类为一簇,说明不同亚型成员具有独立的起源,也说明DGAT家族分化发生在物种分化之前,与Yan等[12]和Rosli等[11]的研究结果一致。同时注意到外类物种蒺藜苜蓿和花生DGAT成员在进化树中区别于其他禾本科作物聚类为同一分支,说明DGAT成员具有一定的种属特异性。基因结构是研究基因家族进化的重要依据。研究[12]发现,不同亚型DGAT成员的外显子数量及基因结构的剪切方式具有明显差异,但同一亚型成员间又高度一致,说明不同DGAT亚型在发生平行进化的同时具有较高保守性,而这种基因结构差异的产生可能是DGAT基因家族进化的一种保守模式。氨基酸序列比对及结构域预测结果说明,不同亚型成员具有特异的结构域,其中包括MBOAT结构域(DGAT1)、DAGAT结构域(DGAT2)和WES_acyltransf结构域(WS/DGAT),也包含其他高度保守的功能区片段,结构域和功能区的差异性说明DOTA不同亚型成员具有不同的生物学特性,与Aymé等[9]和Yan等[12]研究结果一致。基因复制是基因家族在进化过程中产生成员扩增的主要动力。本研究表明,DGAT成员未发生串联复制事件,说明片段复制是DGAT基因家族扩增的主要方式。
表达模式分析是解读基因功能的重要途径。本研究通过构建小麦组织表达图谱,发现不同亚型成员的表达模式存在一定差异。WS/DGAT成员在根中具有较高的表达丰度,但存在部分基因未检测到表达,可能该类基因为环境依赖性转录因子或在某一特定阶段参与表达;而DGAT1和DGAT2成员则在不同组织中均存在高表达基因,说明DGAT1和DGAT2亚型成员中可能存在管家基因。该研究结果与Liu等[5]研究结果不同,判断其原因可能是研究物种不同造成的表达差异,也预示着不同物种具有严格和特异的TAG合成调控方式[24]。顺式作用元件是基因表达调控的关键因素。本研究发现,DGAT成员启动子区域含有大量胁迫反应元件,说明DGAT成员在植株遭遇逆境胁迫过程中具有重要作用。本研究利用已公布转录组数据构建DGAT成员在遭遇干旱、高温和水淹等不同胁迫时的表达模式,发现DGAT成员可广泛响应非生物胁迫,但存在表达特异性。如ZmWSD-1在玉米植株遭遇水淹胁迫时,在敏感品种中高表达,在耐受品种中不表达或表达量较低;ZmDGAT2-3则在不同品种中均未检测到表达量,说明DGAT成员在响应不同非生物胁迫过程中功能分工不同。为验证转录组分析结果,利用实时荧光定量PCR技术,对玉米幼苗进行了ABA、盐和干旱等不同胁迫处理,发现ZmDGATs出现了不同的表达模式,说明这些基因在玉米响应非生物胁迫过程中发挥重要作用,验证了转录组分析的准确性。同时发现,ZmDGAT1-1和ZmDGAT1-2在面对不同胁迫时均上调表达,说明DGAT1家族成员可能整合了多种非生物胁迫信号。
4 结论
本研究通过对主要禾谷类作物进行全基因组扫描,鉴定到玉米7个、水稻7个、小麦26个、高粱9个、谷子10个DGAT家族成员。DGAT可分为4种亚型,不同亚型成员具有独立的起源并平行进化。DGATs广泛参与植株建成过程,不同程度响应非生物胁迫,具有一定功能分工。本研究可为禾谷类作物DGAT基因家族研究奠定基础。
参考文献
Functions of triacylglycerols during plant development and stress
DOI:S0958-1669(17)30085-X
PMID:28987914
[本文引用: 1]

Plant oil in the form of triacylglycerols (TAGs) is a major storage compound used as food, feed and sustainable feedstock for biofuel production. Recent findings suggest that TAGs are more than a carbon and energy reserve in seeds and other storage tissues. In vegetative tissues, TAG metabolism is involved in cell division and expansion, stomatal opening, and membrane lipid remodeling. Moreover, in reproductive tissues, TAGs are important for both organ formation and successful pollination. Here we provide a brief overview of the physiological function and contribution of TAGs during plant development under optimal and varying environmental conditions. These roles of TAGs need to be considered during engineering attempts to further improve TAG content in different tissues.Copyright © 2017 Elsevier Ltd. All rights reserved.
Triacylglycerol Accumulation in photosynthetic cells in plants and algae
Role of DGAT enzymes in triacylglycerol metabolism
DOI:S0003-9861(18)30190-5
PMID:30077544
[本文引用: 1]
The esterification of a fatty acyl moiety to diacylglycerol to form triacylglycerol (TAG) is catalysed by two diacylglycerol O-acyltransferases (DGATs) encoded by genes belonging to two distinct gene families. The enzymes are referred to as DGAT1 and DGAT2 in order of their identification. Both proteins are transmembrane proteins localized in the endoplasmic reticulum. Their membrane topologies are however significantly different. This difference is hypothesized to give the two isozymes different abilities to interact with other proteins and organelles and access to different pools of fatty acids, thereby creating a distinction between the enzymes in terms of their role and contribution to lipid metabolism. DGAT1 is proposed to have dual topology contributing to TAG synthesis on both sides of the ER membrane and esterifying only the pre-formed fatty acids. There is evidence to suggest that DGAT2 translocates to the lipid droplet (LD), associates with other proteins, and synthesizes cytosolic and luminal apolipoprotein B associated LD-TAG from both endogenous and exogenous fatty acids. The aim of this review is to differentiate between the two DGAT enzymes by comparing the genes that encode them, their proposed topologies, the proteins they interact with, and their roles in lipid metabolism.Copyright © 2018 Elsevier Inc. All rights reserved.
High-performance variants of plant diacylglycerol acyltransferase 1 generated by directed evolution provide insights into structure function
DOI:10.1111/tpj.13652 URL [本文引用: 1]
Functional characterization of three novel genes encoding diacylglycerol acyltransferase (DGAT) from oil- rich tubers of cyperus esculentus
DOI:10.1093/pcp/pcz184 URL [本文引用: 2]
Discovery of a new mechanism for regulation of plant triacylglycerol metabolism: The peanut diacylglycerol acyltransferase-1 gene family transcriptome is highly enriched in alternative splicing variants
DOI:S0176-1617(17)30242-0
PMID:29031100
[本文引用: 1]
Triacylglycerols (TAGs) are the most important energy storage form in oilseed crops. Diacylglycerol acyltransferase (DGAT) catalyzes the rate-limiting step of the Kennedy pathway of TAG biosynthesis. To date, little is known about the regulation of DGAT activity in peanut (Arachis hypogaea), an agronomically important oilseed crop that is cultivated in many parts of the world. In this study, seven distinct forms of type 1 DGAT (AhDGAT1.1-AhDGAT1.7) were identified, cloned, and characterized. Comparisons of the nucleotide sequences and gene structures revealed many different splicing variants of AhDGAT1, some of which displayed different organ-specific expression patterns. A representative gene (AhDGAT1.1) was transformed into wild-type tobacco and was shown to increase seed fatty acid (FA) content by 14.7%-20.9%. All seven AhDGAT1s were expressed in TAG-deficient Saccharomyces cerevisiae strain H1246; the five longest AhDGAT1 variants generated high levels of acyltransferase activity and complemented the free fatty acid lethality phenotype in this strain. The alternative splicing that gives rise to AhDGAT1.2 and AhDGAT1.4 creates predicted protein C-terminal truncations. The proteins encoded by these two variants were not active and did not complement the fatty acid sensitivity in H1246. These results were verified by visualization of intracellular lipid droplets using Nile Red staining. Collectively, the results presented here represent the first comprehensive analysis of the peanut DGAT1 gene family, which, unlike in other published plant DGAT1 sequences, shows widespread alternative splicing that may affect the expression patterns and enzyme activities of some members of the gene family.Copyright © 2017. Published by Elsevier GmbH.
AtDGAT2 is a functional acyl-CoA:diacylglycerol acyltransferase and displays different acyl-CoA substrate preferences than AtDGAT1
DOI:10.1016/j.febslet.2013.06.003 URL [本文引用: 1]
Functional characterization of an novel acyl-CoA:diacylglycerol acyltransferase 3-3 (CsDGAT3-3) gene from camelina sativa
DOI:10.1016/j.plantsci.2020.110752 URL [本文引用: 1]
Arabidopsis thaliana DGAT 3 is a [2Fe-2S] protein involved in TAG biosynthesis
DOI:10.1038/s41598-018-35545-7 URL [本文引用: 2]
In silico characterization and expression profiling of the diacylglycerol acyltransferase gene family (DGAT1,DGAT2,DGAT3 and WS/DGAT) from oil palm, Elaeis guineensis
DOI:10.1016/j.plantsci.2018.07.011 URL [本文引用: 2]
Genome-wide characterization and expression profiling of diacylglycerol acyltransferase genes from maize
DOI:10.1139/gen-2018-0029
PMID:30092654
[本文引用: 4]
Diacylglycerol acyltransferase (DGAT) catalyzes the only rate-limiting step in the pathway of plant oil (TAG) biosynthesis and is involved in plant development. In this study, five DGAT family members were identified from maize genome database. Phylogenetic analysis classified the ZmDGATs into type-I, II, and III clusters. Conserved functional domain analysis revealed that the proteins encoded by ZmDGAT1 contained conserved MBOAT domains, while two ZmDGAT2-encoding proteins harbored LPLAT domains. qRT-PCR analysis showed that ZmDGAT genes exhibited very high relative expression in developing seeds, especially at the early stage of seed development. Under various abiotic stress conditions, differential responses of ZmDGAT genes were observed. An overall significant induction of ZmDGAT genes under cold stress in leaves and a quick and strong response to osmotic stresses in roots were highlighted. This study provides useful information for understanding the roles of DGATs in oil accumulation and stress responses in higher plants.
Arabidopsis mutants deficient in diacylglycerol acyltransferase display increased sensitivity to abscisic acid,sugars,and osmotic stress during germination and seedling development
DOI:10.1104/pp.006122 URL [本文引用: 1]
Directed evolution of acyl-CoA:diacylglycerol acyltransferase:development and characterization of Brassica napus DGAT1 mutagenized libraries
DOI:10.1016/j.plaphy.2008.12.019 URL [本文引用: 1]
Oil crops for the future
DOI:S1369-5266(19)30118-9
PMID:31982290
[本文引用: 1]
Agriculture faces enormous challenges including the need to substantially increase productivity, reduce environmental footprint, and deliver renewable alternatives that are being addressed by developing new oil crops for the future. The efforts include domestication of Lepidium spp. using genomics-aided breeding as a cold hardy perennial high-yielding oil crop that provides substantial environmental benefits, expands the geography for oil crops, and improves farmers' economy. In addition, genetic engineering in Crambe abyssinica may lead to a dedicated industrial oil crop to replace fossil oil. Redirection of photosynthates from starch to oil in plant tubers and cereal endosperm also provides a path for enhancing oil production to meet the growing demands for food, fuel, and biomaterials. Insect pheromone components are produced in seed oil plants in a cost-effective and environmentally friendly pest management replacing synthetically produced pheromones. Autophagy is explored for increasing crop fitness and oil accumulation using genetic engineering in Arabidopsis.Copyright © 2019 Elsevier Ltd. All rights reserved.
Non-edible vegetable oil-based feedstocks capable of bio-lubricant production for automotive sector applications-a review
DOI:10.1007/s11356-019-05000-9 URL [本文引用: 1]
The third revolution in sequencing technology
DOI:S0168-9525(18)30096-9
PMID:29941292
[本文引用: 1]
Forty years ago the advent of Sanger sequencing was revolutionary as it allowed complete genome sequences to be deciphered for the first time. A second revolution came when next-generation sequencing (NGS) technologies appeared, which made genome sequencing much cheaper and faster. However, NGS methods have several drawbacks and pitfalls, most notably their short reads. Recently, third-generation/long-read methods appeared, which can produce genome assemblies of unprecedented quality. Moreover, these technologies can directly detect epigenetic modifications on native DNA and allow whole-transcript sequencing without the need for assembly. This marks the third revolution in sequencing technology. Here we review and compare the various long-read methods. We discuss their applications and their respective strengths and weaknesses and provide future perspectives.Copyright © 2018 Elsevier Ltd. All rights reserved.
TBtools: an integrative toolkit developed for interactive analyses of big biological data
DOI:S1674-2052(20)30187-8
PMID:32585190
[本文引用: 1]
The rapid development of high-throughput sequencing techniques has led biology into the big-data era. Data analyses using various bioinformatics tools rely on programming and command-line environments, which are challenging and time-consuming for most wet-lab biologists. Here, we present TBtools (a Toolkit for Biologists integrating various biological data-handling tools), a stand-alone software with a user-friendly interface. The toolkit incorporates over 130 functions, which are designed to meet the increasing demand for big-data analyses, ranging from bulk sequence processing to interactive data visualization. A wide variety of graphs can be prepared in TBtools using a new plotting engine ("JIGplot") developed to maximize their interactive ability; this engine allows quick point-and-click modification of almost every graphic feature. TBtools is platform-independent software that can be run under all operating systems with Java Runtime Environment 1.6 or newer. It is freely available to non-commercial users at https://github.com/CJ-Chen/TBtools/releases.Copyright © 2020 The Author. Published by Elsevier Inc. All rights reserved.
Acyl-CoA:diacylglycerol acyltransferase:molecular biology,biochemistry and biotechnology
DOI:10.1016/j.plipres.2012.06.001 URL [本文引用: 1]
Structure-function analysis of diacylglycerol acyltransferase sequences from 70 organisms
DOI:10.1186/1756-0500-4-249
PMID:21777418
[本文引用: 1]
Diacylglycerol acyltransferase families (DGATs) catalyze the final and rate-limiting step of triacylglycerol (TAG) biosynthesis in eukaryotic organisms. Understanding the roles of DGATs will help to create transgenic plants with value-added properties and provide clues for therapeutic intervention for obesity and related diseases. The objective of this analysis was to identify conserved sequence motifs and amino acid residues for better understanding of the structure-function relationship of these important enzymes.117 DGAT sequences from 70 organisms including plants, animals, fungi and human are obtained from database search using tung tree DGATs. Phylogenetic analysis separates these proteins into DGAT1 and DGAT2 subfamilies. These DGATs are integral membrane proteins with more than 40% of the total amino acid residues being hydrophobic. They have similar properties and amino acid composition except that DGAT1s are approximately 20 kDa larger than DGAT2s. DGAT1s and DGAT2s have 41 and 16 completely conserved amino acid residues, respectively, although only two of them are shared by all DGATs. These residues are distributed in 7 and 6 sequence blocks for DGAT1s and DGAT2s, respectively, and located at the carboxyl termini, suggesting the location of the catalytic domains. These conserved sequence blocks do not contain the putative neutral lipid-binding domain, mitochondrial targeting signal, or ER retrieval motif. The importance of conserved residues has been demonstrated by site-directed and natural mutants.This study has identified conserved sequence motifs and amino acid residues in all 117 DGATs and the two subfamilies. None of the completely conserved residues in DGAT1s and DGAT2s is present in recently reported isoforms in the multiple sequences alignment, raising an important question how proteins with completely different amino acid sequences could perform the same biochemical reaction. The sequence analysis should facilitate studying the structure-function relationship of DGATs with the ultimate goal to identify critical amino acid residues for engineering superb enzymes in metabolic engineering and selecting enzyme inhibitors in therapeutic application for obesity and related diseases.
Gene duplication and evolution in recurring polyploidization-diploidization cycles in plants
DOI:10.1186/s13059-019-1650-2
PMID:30791939
[本文引用: 1]
The sharp increase of plant genome and transcriptome data provide valuable resources to investigate evolutionary consequences of gene duplication in a range of taxa, and unravel common principles underlying duplicate gene retention.We survey 141 sequenced plant genomes to elucidate consequences of gene and genome duplication, processes central to the evolution of biodiversity. We develop a pipeline named DupGen_finder to identify different modes of gene duplication in plants. Genes derived from whole-genome, tandem, proximal, transposed, or dispersed duplication differ in abundance, selection pressure, expression divergence, and gene conversion rate among genomes. The number of WGD-derived duplicate genes decreases exponentially with increasing age of duplication events-transposed duplication- and dispersed duplication-derived genes declined in parallel. In contrast, the frequency of tandem and proximal duplications showed no significant decrease over time, providing a continuous supply of variants available for adaptation to continuously changing environments. Moreover, tandem and proximal duplicates experienced stronger selective pressure than genes formed by other modes and evolved toward biased functional roles involved in plant self-defense. The rate of gene conversion among WGD-derived gene pairs declined over time, peaking shortly after polyploidization. To provide a platform for accessing duplicated gene pairs in different plants, we constructed the Plant Duplicate Gene Database.We identify a comprehensive landscape of different modes of gene duplication across the plant kingdom by comparing 141 genomes, which provides a solid foundation for further investigation of the dynamic evolution of duplicate genes.
Triacylglycerol metabolism,function,and accumulation in plant vegetative tissues
DOI:10.1146/annurev-arplant-043015-111641 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}