


元素硒(Se)于1817年首次被瑞典科学家Jons Jakob Berzelius发现[1],并一度被认为是有毒物质。直至1957年,硒的营养作用才被挖掘出来。人体中多种重要的生理作用受硒蛋白调控,如人体免疫力、维持氧化还原平衡、预防心脑血管疾病和调节降低肿瘤风险等,已知硒蛋白有25种,硒是人体必需的微量元素之一。全世界有40多个国家和地区存在缺硒现象,大约有10亿人口受到硒缺乏的影响[2];中国2/3人口硒摄入量不足,全国不同程度缺硒现象占72%。目前,通过食物补硒是提高人体摄硒量的最佳途径,食物中硒含量决定着食物链中的硒水平,因此,探究植物对硒的吸收方式以及硒在植物体内的转运及代谢机制,对富硒植物的创新性生产具有重要的指导意义。
硒是一把双刃剑,低浓度的硒对植物生理功能的实现有重要意义,可提高种子发芽率,增强光合作用,增加作物产量,促进植物生长,以及提高植物抵御多种逆境的能力[3⇓-5];高浓度的硒反而使有些植物出现中毒症状,损伤叶绿体膜的完整性,导致基粒结构解体,抑制植株生长及各项生理活动[6]。至今尚未证明硒是植物的必需元素,但大部分研究显示,适宜浓度的硒在植物生长、作物品质以及植物抗逆性等方面起有益作用。根据植物在其自然栖息地积累硒的能力,可划分为非积累型(<100mg Se/kg干重)、积累型(100~1000mg Se/kg干重)和超积累型(>1000mg Se/kg干重),其中十字花科作物对硒的积累能力最强,其次是豆科,谷类最低。谷类中小麦对硒积聚最多[7]。因此,了解植物对硒的吸收、转运和代谢机制,有助于科学指导富硒植物产品的开发与利用,也有益于拓展硒在农业生态领域安全的应用。
1 环境中的硒
1.1 环境中硒的循环途径
自然界中的硒具有一套复杂的生物化学循环系统,通常以负二价硒离子Se(−II)、正四价硒离子Se(IV)、正六价硒离子Se(VI)和单价硒Se(0)4种价态存在于不同的无机和有机硒化物中[8]。
大气环境中的硒主要由人为排放的硒和自然来源的硒构成,以气态和颗粒态的形式存在,包括气态有机化合物二甲基硒(DMSe)和二甲基二硒(DMDSe),气态无机化合物(SeO2、H2Se)和附着于颗粒表面的元素硒Se(0)。硒矿石的开采、提炼以及煤炭的燃烧会生成SeO2排入大气,约占大气硒含量的40%[11];余下60%多是自然来源,包括火山喷发带来的H2Se、部分有机物的挥发,以及来自海浪、灰尘的颗粒物表面所附着的Se(0)[12]。土壤中的硒则主要源自硒矿石和煤矿等富硒资源的自然释放,但在英国西部和中国东南部等地区,大气中硒的沉降作用也是土壤硒的重要来源[13]。土壤硒含量主要由地质决定,同时受到土壤类型、有机质和降雨量等条件影响,使全球土壤硒分布具有空间异质性[14]。自然环境中土壤硒含量普遍在0.01~ 2.0mg/kg,世界平均硒含量为0.4mg/kg[15]。就我国土壤而言,缺硒面积高达72%,土壤硒含量普遍在0.2~3.0mg/kg范围,既有严重缺硒地区,也存在硒污染地区[16]。自然水体中硒含量较低,一般处于10μg/L以下[17],多数以硒酸盐和亚硒酸盐的形式存在[18],主要来源于电子工业、饲料、养殖业和医药行业等人类生产生活活动,也有部分来源于土壤硒。
微生物途径是环境硒循环的重要途径,在修复硒污染的过程中具有重要意义。研究[19]表明,微生物相较于动植物能对含有高浓度Se(IV)和Se(VI)的生存环境产生强耐受性,同时还可以经由众多途径对硒的形态进行转化。微生物循环中的硒转化途径包括氧化途径、异化还原途径和同化还原途径,还包含脱盐、甲基化和去甲基化等其他途径(图1)。在一系列转化途径中,微生物不仅可以促进将高浓度Se(IV)和Se(VI)转化为毒性较低且不具有可溶性的纳米硒(Selenium nanoparticles,SeNPs),也可以将其同化还原为硒蛋白,或经过甲基化途径生成高挥发性的甲基硒化物[17,20-21],一些低价态的硒还可以通过微生物氧化为高价态的硒,因此在降低硒毒害地区的危害性中具有广阔的应用前景。
图1

1.2 硒在土壤中的存在形态及有效性
硒以Se(−II)、Se(IV)、Se(VI)和Se(0)多种价态存在于土壤中,不同形态硒在土壤中的分布与土壤的有机质、氧化还原电位(Eh)、土壤类型、螯合物、pH和气候等因素密切相关,不同价态的硒具有不同的有效性。
其中,Se(IV)和Se(VI)是易被生物利用的2种主要硒形态,前者占土壤硒总量的40%以上,后者不超过10%[27]。Se(VI)在含氧、干燥、偏碱性[(pE+pH)>15]的土壤中含量较多,易被植物吸收利用;Se(IV)则主要存在于缺氧、潮湿、中性到酸性[7.5<(pE+pH)<15]的土壤中,但易被铁铝氧化物等矿物胶体吸附和固定,显著降低亚硒酸盐的移动性和溶解性,进而在一定程度上降低硒的有效性[28]。Se(0)难以被植物利用,在条件适宜时,可经水解或微生物氧化作用转化为对植物有效的硒形态;Se(−II)一般难以溶解迁移,易形成沉积物在土壤中积累,生物有效性较低,一些可溶性硒可被植物吸收利用,但是需要通过风化作用缓慢释放[29]。
土壤中的有机硒是一种来自于植物和微生物同化还原过程产生潜在的可利用硒,包括硒代半胱氨酸(SeCys)、硒代蛋氨酸(SeMet)、甲基硒代半胱氨酸(MeSeCys)等。研究[30]表明,植物可能通过细胞膜上的氨基酸转运蛋白和脯氨酸通透酶途径等方式直接吸收这几种形态的有机硒。除了上述具有明确化学结构的有机硒化合物,土壤中还存在有机质结合态硒,可通过有机质直接络合吸附Se含氧阴离子和有机质―金属桥键―Se三元络合物2种方式形成,是补充土壤有机硒的重要途径。有机结合态硒是土壤Se迁移转化过程的潜在“硒池”,主要成分为胡敏酸结合态硒(HA-Se)和富里酸结合态硒(FA-Se),它的生物有效性取决于矿化和还原作用(矿化利用和还原固定)的相对强度[31]。例如,可被植物吸收利用的Se(IV)来自于易被矿化分解的FA-Se、Se(VI)和低分子有机硒化合物,生物有效性高于HA-Se。土壤中FA-Se基本占有机结合态硒60%以上,且FA-Se与HA-Se之间也存在着相互转化[32]。
2 植物对硒的吸收和转运机制
植物可通过根系和叶片吸收硒,但吸收方式受硒形态及植物部位的影响。土壤或溶液中的无机态硒(SeO42-、SeO32-、HSeO3-、H2SeO3)和少量小分子有机硒(SeCys、SeMet)能够被植物根系吸收,但胶体硒和金属硒化物不能被植物的根系吸收。植物叶片则可以吸收无机硒、有机硒乃至纳米单质硒[33]。
2.1 植物根部对硒酸盐的吸收与转运
硒和硫是元素周期表中同主族元素,具有非常相似的生物化学性质,除超积累型植物外,大部分植物并不能有效区分二者。因此,植物根系可通过高亲和力的硫酸盐转运蛋白(SULTR)以主动耗能的形式吸收土壤中的硒酸盐。植物中有4类H+/硫酸盐同向转运蛋白与硒酸盐吸收转运相关,在根部、维管束系统和叶片等不同部位发挥相应的吸收或运输功能(图2)。
图2

图2
植物对硒酸盐的吸收和转运机制
Fig.2
The absorption and transport mechanism of selenate by plants
第1类硫酸盐转运子(SULTR1;1、SULTR1;2和SULTR1;3)都是具有高亲和力的转运蛋白。SULTR1;1和SULTR1;2主要在根部的表皮中表达,在植物从根际土壤吸收硒酸盐的过程中发挥着重要的作用。其中,SULTR1;2对硫酸盐和硒酸盐的吸收贡献较大,而SULTR1;1对2种离子的吸收程度较低[34]。这可能与SULTR1;1和SULTR1;2的表达水平有关,后者在植物根中的表达量比前者高出约10倍[35]。Elie等[36]在研究拟南芥硫酸盐转运蛋白家族13个突变基因时发现,只有SULTR1;2基因的突变导致了对拟南芥的硒酸盐抗性,说明SULTR1;2是植物体内唯一调控根部吸收硒酸盐的硫酸盐转运体。SULTR1;3则定位于根和茎的韧皮部中,参与硒酸盐在韧皮部中的吸收与转运[37]。Boldrin等[38]研究表明,在硒酸盐处理的小麦中SULTR1;3的表达量明显上调,但该转运蛋白的准确功能仍需进一步的研究。
第2类转运子是低亲和力转运蛋白(SULTR2;1和SULTR2;2),主要负责硒的迁移转运,分布在叶、根和维管束系统等器官。低亲和力的硫酸盐转运蛋白SULTR2;1和SULTR2;2将吸收到植物体内的硒酸盐运输到木质部,并转移到植物的地上部分。El等[39]研究发现,高表达的SULTR2;1增强了沙漠王羽(Stanleya pinnata)对硒酸盐吸收转运的特异性,且不受外界环境中硫酸盐含量的影响。
第3类硫酸盐转运子(SULTR3;1和SULTR3;5)定位于叶绿体膜上。硒酸盐主要通过SULTR3;1跨膜进入叶绿体,叶绿体中硒酸盐的含量随着SULTR3;1的缺失而降低。SULTR3;5不仅实现硒酸盐在木质部的长距离运输,还可以进一步将硒酸盐转运至叶绿体中[40]。此外,SULTR3;5具有可以与其他类别的转运蛋白协同转运硒酸盐的能力。Kataoka等[41]研究发现,SULTR3;5可以显著增强SULTR2;1的活性,提高拟南芥对硒酸盐的吸收能力;在敲除SULTR3;5同时SULTR2;1过表达的植株中,硒酸盐从根到茎的运输明显减少,这就表明SULTR3;5和SULTR2;1在将硒酸盐由根部转运到木质部的过程中具有协同作用。
除上述硫酸盐转运蛋白外,硒酸盐还可以利用电化学梯度进行转运。比如,液泡相对于细胞质而言带正电,硒酸根可经由阴离子通道沿其电化学梯度向下被动运输进入液泡[46];叶绿体基质中的总电荷相对于细胞质溶胶显负电,硒酸盐可沿其电化学梯度向下移出叶绿体。
2.2 植物根部对亚硒酸盐的吸收与转运
关于植物对硒酸盐的吸收途径研究甚多,但关于亚硒酸盐的吸收方式探究相对较少。早期的研究[47]表明,植物根系吸收亚硒酸盐可能通过被动扩散实现,因为尚未发现相关的膜介导转运。随着相关研究的不断深入,Li等[48]在研究小麦(Triticum aestivum)对无机硒的吸收机制时发现,亚硒酸盐的吸收效率受到代谢抑制剂羰基氰化物间氯苯腙的影响,推测植物对亚硒酸盐的吸收过程是一种依赖于代谢的主动吸收过程,且受部分磷转运体的介导。随后,Song等[49]和Zhang等[50]分别在烟草(Nicotiana tabacum)和水稻(Oryza sativa)中证实,无机磷酸盐转运蛋白可以在根部吸收亚硒酸盐,且特定磷酸盐转运蛋白的过表达会显著提高植物对亚硒酸盐的吸收量(比如,OsPT8过表达的烟草可积累更多的硒)。亚硒酸盐的吸收载体不仅仅只有磷酸盐转运蛋白,Zhao等[51]发现,水稻吸收亚硒酸盐是通过硅元素的转运载体OsNIP2;1,并且被环境pH影响。由此可见,植物对亚硒酸盐的吸收途径复杂且多样化,远不及硒酸盐的研究深入,因此有待从生理及分子角度继续挖掘。
值得注意的是,植物根系对亚硒酸盐有较强的亲和力,在根系中会固定住大部分的亚硒酸盐,只有少部分转化为有机硒化物(如硒代蛋氨酸SeMet、硒甲基半胱氨酸MeSeCys等)经由木质部转运至地上部。Jiang等[52]对荞麦(Fagopyrum esculentum M.)的研究证实,荞麦根部在施加亚硒酸钠后积累了大量的硒。但也有研究[53]表明,胡萝卜施加亚硒酸盐后表现出根部硒含量最低,茎与叶中硒的含量相对较高的现象。此外,Li等[48]研究也指出,将硒酸盐和亚硒酸盐同时施加到植物中,硒酸盐从植物地下部到茎、叶的转运可能受到亚硒酸盐的抑制。因此,针对不同作物对硒酸盐和亚硒酸盐的吸收和转运效率的差别,以及硒酸盐和亚硒酸盐的转运是否会相互影响仍需进一步研究。
2.3 植物根部对有机硒的吸收与转运
李玉梅等[55]证明有机硒可以提升作物对硒的富集能力,在满足作物生长的营养需求之外,提升可食用部分的硒含量。Kikkert等[56]发现,水培试验中小麦和油菜对有机硒的吸收效率比无机硒高20~100倍,且施加SeMet的植物体内检测出的硒浓度最高,推测有机硒(尤其是SeMet)较容易被植物吸收并转移到地上部。不仅如此,当对植物同时供应外源SeMet和蛋氨酸时,植物对SeMet的吸收受蛋氨酸的影响较大。早期研究[57]表明,植物可能通过植物根系蛋氨酸转运蛋白吸收SeMet。邓坤[58]在研究水稻对SeMet的吸收转运机制时发现,呼吸抑制剂间氯苯腙和2,4-二硝基苯酚能够显著抑制水稻根系对SeMet的吸收,推测SeMet的吸收与植物能量代谢有关。随后,王琪[59]研究表明,水通道蛋白抑制剂AgNO3作用于水稻根系对SeMet吸收的抑制效果高达93.1%,推测有机硒可能会利用水通道蛋白进入植物根内。王琪[59]也发现添加SeMet和硒代蛋氨酸氧化物这些外源有机硒后,根系和地上部主要以有机硒MeSeCys和SeMet形式存在,且水稻根系对SeMet的吸收速率显著高于其他形态的硒处理。此外,水稻肽转运蛋白(PTR)家族成员NRT1.1B和硝酸盐转运蛋白CHR1也被证实具有转运SeMet的功能[60]。
2.4 植物叶片对硒的吸收与转运
叶片本身不是用来吸收养分的营养器官,也没有进化出吸收硒元素的特异性结构。但在长期的农业实际生产中,喷施硒肥可以显著提高植物体内的硒含量,这表明硒可以通过叶片进入植物体内。尽管目前关于叶片吸收硒的研究较少,但可以从叶片的结构推测硒吸收的潜在途径可能与角质层、气孔和毛状体等表皮结构有关[61]。
大多数植物叶片的表面覆盖着一层角质层,它是叶片表面积占比最大以及与外源物质接触面积最大的区域[62]。作为一种物理屏障,角质层能保护叶片免受水分损失、辐射及病原体的伤害。此外,角质层膜对有机态和无机态的粒子具有渗透性,如水、无机盐离子和有机分子等,因此硒通过角质层进入叶片可能是一个扩散解吸的过程。
3 植物对硒的转化与代谢
植物以硒酸盐、亚硒酸盐或有机硒形式吸收利用硒。进入植物体内的硒转化为各种有机形式主要通过代谢来实现转化。硒在根部通过硒酸盐的还原、硒代氨基酸的合成和挥发性硒的形成代谢途径进入植物。
3.1 植物对硒酸盐的转化与代谢
硒酸盐的转化和代谢主要发生在叶细胞的细胞液和质体中的,已有众多研究[65-66]表明,硒酸盐进入质体的途径与硫酸盐类似,均由SULTRs介导(图3)。质体中,在ATP硫酸化酶(ATPS)的作用下,硒酸盐首先被活化成5′-磷酸硒腺苷(APSe),然后在5′-磷硫酸腺苷还原酶(APR)的作用下转化为亚硒酸盐[67]。Sors等[68]对拟南芥的研究表明,过量的ATPS还原酶和APR都可以增加植物中亚硒酸盐的比例。然后,亚硒酸盐被还原成Se2-是在亚硫酸还原酶(SiR)的作用下实现的。但有研究[69]表明,SiR的敲除或抑制并不能影响亚硒酸盐的还原。还有研究[70]认为亚硒酸盐可以通过与谷胱甘肽的相互作用而被还原。因而,此处的还原机制仍有待研究。在半胱氨酸合成酶(CSase)的作用下,Se2-进一步通过O-乙酰丝氨酸巯基裂解酶(OASTL)和丝氨酸乙酰转移酶(SAT)建立可逆性复合物,生成SeCys[71]。SeCys产生后,硒的代谢分为3条路径,第1个是SeCys在硒代半胱氨酸裂解酶(SL)的作用下直接释放出元素硒;第2个是在硒代半胱氨酸甲基转移酶(SMT)的作用下,SeCys转化为SeMSeCys,接着转化为可挥发的二甲基二硒醚(DMDse);第3个则是SeCys向SeMet的转化[28]。
图3

在SeCys向SeMet转化的过程中,首先,胱硫醚γ-合成酶(CγS)可以将SeCys合成为硒代胱硫醚(SeCysta),在胱硫醚-β-裂解酶(CβL)的作用下,SeCysta分裂形成硒代高半胱氨酸(SeHCys),之后SeHCys在蛋氨酸合成酶(MTR)的作用下转化成SeMet[72]。
但这一步的转化只是侧面的推断,对哺乳动物的研究[73]中,MTR作为催化剂促进了SeMet的生成,因此猜测植物中的MTR具有相似的功能,可以作用于SeHCys促进SeMet生成。在蛋氨酸转移酶(MMT)的作用下,SeMet转化成了硒甲基硒代蛋氨酸(SeMSeMet),而后SeMSeMet转化为二甲基硒化物(DMSe)这种具有挥发性的化合物[74]。SeMSeMet向DMSe转化可能有2种方式,在一些植物中,SeMSeMet被甲基甲硫氨酸水解酶(MMH)直接分解从而形成DMSe[75];另外一种方式猜测是SeMSeMet生成DMSe的过程中存在中间产物,这类中间产物可能是一种通过未知过程快速将SeMSeMet转化为挥发性DMSe的“中转站”[76]。
3.2 植物对亚硒酸盐和有机硒的转化与代谢
目前关于亚硒酸盐和有机硒代谢的研究较少,主要是由于植物吸收的亚硒酸盐大部分积累在根部,并很快地转化为有机硒,进而造成木质部亚硒酸盐含量很低[48]。由于根中积累的有机硒主要为非水溶性硒化物,所以木质部和叶片中直接从外界吸收的亚硒酸盐含量较少,相关的代谢途径仍有待研究。植物吸收有机硒的来源有两部分,一是来自于根系直接从土壤中吸收的硒代半胱氨酸和硒代蛋氨酸,另外一部分来自于根部亚硒酸盐转化而来的有机硒,这部分有机硒可能直接参与到硒酸盐的代谢途径当中。
4 展望
4.1 生物强化和植物修复
研究[82]发现,微生物将硒酸盐和亚硒酸盐异化还原为Se(0)是硒污染环境的主要修复手段,期间产生的纳米硒,毒性微弱近乎无毒。如今,硒酸盐及亚硒酸盐的异化还原过程是目前针对如何减少硒污染的主要研究内容,同时针对毒性较低的硒形态去提高硒的生物利用度,也有利于人体合理补硒。
4.2 硒的吸收转运和代谢机制的进一步研究
对于植物修复和生物强化的改进需要对硒的吸收转运代谢途径具有透彻的了解。植物中没有硒特异性途径,似乎不需要硒,但实际上硒通过硫转运蛋白被转运,并通过硫代谢酶被代谢,因此了解硒的运输和代谢及理解硫的运输和代谢过程,可以应用于提高硒的吸收和同化,使其在特定的物种中变成无毒形式,可以用于植物修复和生物强化。
关于硒酸盐的吸收转运和代谢的过程研究较多,机制较为清晰(图4)。一般认为硒和硫具有相似的特性,植物主要通过硫酸盐转运体吸收硒酸盐。为了进一步比较不同硒转运体的转运能力和特异性,可以通过离子通道的鉴定分析方法建立硒酸盐转运体的标准鉴定方法,这也为进一步比较同种和不同种不同转运载体的运输特性和能力差异奠定基础。
图4
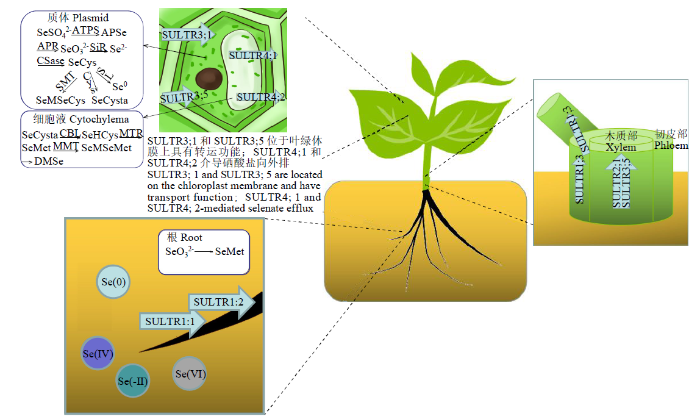
图4
植物中硒酸盐的吸收转运及代谢示意图
Fig.4
Schematic diagram of selenate absorption, transport and metabolism in plants
亚硒酸盐的吸收和转运机理有待进一步研究。结合已有研究结果,推测亚硒酸盐的吸收和转运途径为H2SeO3/SeO2转运蛋白途径、HSeO3-和SeO32- 一个转运蛋白途径和一个外排途径。为了获取在植物内亚硒酸盐外排蛋白的基础数据,可以进一步分析亚硒酸盐在植物体内的吸收转运载体及代谢,建立亚硒酸盐转运鉴定的标准方法。
现阶段关于植物吸收有机硒的研究较少,无机硒比有机硒有更高的毒性和生物利用度,因此对于有机硒的研究是生物强化的一条重要途径。
参考文献
The discovery of the elements. VI. Tellurium and selenium
DOI:10.1021/ed009p474 URL [本文引用: 1]
The fascinating facets of plant selenium accumulation-biochemistry,physiology,evolution and ecology
DOI:10.1111/nph.14378
PMID:27991670
[本文引用: 1]

Contents 1582 I. 1582 II. 1583 III. 1588 IV. 1590 V. 1592 1592 References 1592 SUMMARY: The importance of selenium (Se) for medicine, industry and the environment is increasingly apparent. Se is essential for many species, including humans, but toxic at elevated concentrations. Plant Se accumulation and volatilization may be applied in crop biofortification and phytoremediation. Topics covered here include beneficial and toxic effects of Se on plants, mechanisms of Se accumulation and tolerance in plants and algae, Se hyperaccumulation, and ecological and evolutionary aspects of these processes. Plant species differ in the concentration and forms of Se accumulated, Se partitioning at the whole-plant and tissue levels, and the capacity to distinguish Se from sulfur. Mechanisms of Se hyperaccumulation and its adaptive significance appear to involve constitutive up-regulation of sulfate/selenate uptake and assimilation, associated with elevated concentrations of defense-related hormones. Hyperaccumulation has evolved independently in at least three plant families, probably as an elemental defense mechanism and perhaps mediating elemental allelopathy. Elevated plant Se protects plants from generalist herbivores and pathogens, but also gives rise to the evolution of Se-resistant specialists. Plant Se accumulation affects ecological interactions with herbivores, pollinators, neighboring plants, and microbes. Hyperaccumulation tends to negatively affect Se-sensitive ecological partners while facilitating Se-resistant partners, potentially affecting species composition and Se cycling in seleniferous ecosystems.© 2016 The Authors. New Phytologist © 2016 New Phytologist Trust.
Influence of selenium biofortification on the bioactive compounds and antioxidant activity of wheat microgreen extract
DOI:10.1016/j.foodchem.2019.125763 URL [本文引用: 1]
Evaluation of cytotoxicity,biochemical profile and yield components of groundnut plants treated with nano-selenium
DOI:10.1016/j.btre.2019.e00377 URL [本文引用: 1]
Underlying mechanisms responsible for restriction of uptake and translocation of heavy metals (metalloids) by selenium via root application in plants
DOI:10.1016/j.jhazmat.2020.123570 URL [本文引用: 1]
An overview of selenium uptake,metabolism,and toxicity in plants
Metal hyperaccumulator plants:A review of the ecology and physiology of a biological resource for phytoremediation of metal-polluted soils
Delineation of the molecular basis for selenium-induced growth arrest in human prostate cancer cells by oligonucleotide array
Despite the growing interest in selenium intervention of prostate cancer in humans, scanty information is currently available on the molecular mechanism of selenium action. Our past research indicated that methylseleninic acid (MSA) is an excellent reagent for investigating the anticancer effect of selenium in vitro. The present study was designed to examine the cellular and molecular effects of MSA in PC-3 human prostate cancer cells. After exposure to physiological concentrations of MSA, these cells exhibited a dose- and time-dependent inhibition of growth. MSA retarded cell cycle progression at multiple transition points without changing the proportion of cells in different phases of the cell cycle. Flow cytometric analysis of annexin V- and propidium iodide-labeled cells showed a marked induction of apoptosis by MSA. Array analysis with the Affymetrix human genome U95A chip was then applied to profile the gene expression changes that might mediate the effects of selenium. Gene profiling was done in a time course experiment (at 12, 24, 36, and 48 h) using synchronized cells. A large number of potential selenium-responsive genes with diverse biological functions were identified. These genes fell into 12 clusters of distinct kinetics pattern of modulation by MSA. The expression changes of 10 genes known to be critically involved in cell cycle regulation were selected for verification by Western analysis to determine the reliability of the array data. An agreement rate of 70% was obtained based on these confirmation experiments. The array data enabled us to focus on the role of potential key genes (e.g., GADD153, CHK2, p21(WAF1), cyclin A, CDK1, and DHFR) that might be targets of MSA in impeding cell cycle progression. The data also provide valuable insights into novel biological effects of selenium, such as inhibition of cell invasion, DNA repair, and stimulation of transforming growth factor beta signaling. The present study demonstrates the utility of a genome-wide analysis to elucidate the mechanism of selenium chemoprevention.
Arsenic and selenium
Plants,selenium and human health
DOI:10.1016/S1369-5266(03)00030-X URL [本文引用: 1]
Selenium deficiency and toxicity in the environment
Reviews on atmospheric selenium:emissions,speciation and fate
DOI:10.1016/j.atmosenv.2007.07.035 URL [本文引用: 1]
Selenium cycling across soil- plant-atmosphere interfaces:A critical review
DOI:10.3390/nu7064199
PMID:26035246
[本文引用: 1]
Selenium (Se) is an essential element for humans and animals, which occurs ubiquitously in the environment. It is present in trace amounts in both organic and inorganic forms in marine and freshwater systems, soils, biomass and in the atmosphere. Low Se levels in certain terrestrial environments have resulted in Se deficiency in humans, while elevated Se levels in waters and soils can be toxic and result in the death of aquatic wildlife and other animals. Human dietary Se intake is largely governed by Se concentrations in plants, which are controlled by root uptake of Se as a function of soil Se concentrations, speciation and bioavailability. In addition, plants and microorganisms can biomethylate Se, which can result in a loss of Se to the atmosphere. The mobilization of Se across soil-plant-atmosphere interfaces is thus of crucial importance for human Se status. This review gives an overview of current knowledge on Se cycling with a specific focus on soil-plant-atmosphere interfaces. Sources, speciation and mobility of Se in soils and plants will be discussed as well as Se hyperaccumulation by plants, biofortification and biomethylation. Future research on Se cycling in the environment is essential to minimize the adverse health effects associated with unsafe environmental Se levels.
Selenium accumulation by plants
DOI:10.1093/aob/mcv180
PMID:26718221
[本文引用: 1]
Selenium (Se) is an essential mineral element for animals and humans, which they acquire largely from plants. The Se concentration in edible plants is determined by the Se phytoavailability in soils. Selenium is not an essential element for plants, but excessive Se can be toxic. Thus, soil Se phytoavailability determines the ecology of plants. Most plants cannot grow on seleniferous soils. Most plants that grow on seleniferous soils accumulate <100 mg Se kg(-1) dry matter and cannot tolerate greater tissue Se concentrations. However, some plant species have evolved tolerance to Se, and commonly accumulate tissue Se concentrations >100 mg Se kg(-1) dry matter. These plants are considered to be Se accumulators. Some species can even accumulate Se concentrations of 1000-15 000 mg Se kg(-1 )dry matter and are called Se hyperaccumulators.This article provides an overview of Se uptake, translocation and metabolism in plants and highlights the possible genetic basis of differences in these between and within plant species. The review focuses initially on adaptations allowing plants to tolerate large Se concentrations in their tissues and the evolutionary origin of species that hyperaccumulate Se. It then describes the variation in tissue Se concentrations between and within angiosperm species and identifies genes encoding enzymes limiting the rates of incorporation of Se into organic compounds and chromosomal loci that might enable the development of crops with greater Se concentrations in their edible portions. Finally, it discusses transgenic approaches enabling plants to tolerate greater Se concentrations in the rhizosphere and in their tissues.The trait of Se hyperaccumulation has evolved several times in separate angiosperm clades. The ability to tolerate large tissue Se concentrations is primarily related to the ability to divert Se away from the accumulation of selenocysteine and selenomethionine, which might be incorporated into non-functional proteins, through the synthesis of less toxic Se metabilites. There is potential to breed or select crops with greater Se concentrations in their edible tissues, which might be used to increase dietary Se intakes of animals and humans.© The Author 2015. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Selenium distribution in the Chinese environment and its relationship with human health:A review
DOI:10.1016/j.envint.2017.12.035 URL [本文引用: 1]
Dissolved constituents including selenium in waters in the vicinity of kesterson national wildlife refuge and the west grassland,fresno and merced counties,California
DOI:10.1021/tx400428f URL [本文引用: 2]
Selenium speciation of soil/sediment determined with sequential extractions and hydride generation atomic absorption spectrophotometry
DOI:10.1021/es960214+ URL [本文引用: 1]
Selenium:environmental significance,pollution,and biological treatment technologies
DOI:10.1016/j.biotechadv.2016.05.005 URL [本文引用: 1]
Selenium biomineralization for biotechnological applications
DOI:10.1016/j.tibtech.2015.03.004 URL [本文引用: 1]
Selenium uptake by sulfur-accumulating bacteria
DOI:10.1016/0016-7037(96)00221-9 URL [本文引用: 1]
Similarities between the abiotic reduction of selenite with glutathione and the dissimilatory reaction mediated by Rhodospirillum rubrum and Escherichia coli
DOI:10.1074/jbc.M405887200 URL [本文引用: 1]
Selenite reduction by Shewanella oneidensis MR-1 is mediated by fumarate reductase in periplasm
DOI:10.1038/srep03735 [本文引用: 1]
Selenium metabolism in plants
Selenium in soil- microbe-plant systems:sources,distribution,toxicity,tolerance,and detoxification
植物对土壤硒的吸收转化研究进展
DOI:10.19586/j.2095-2341.2017.0104
[本文引用: 1]
硒是人类、动物和某些微生物必需的微量元素。植物因其具有吸收和转化土壤硒的能力而成为人类饮食补硒的主要来源。提高植物的富硒水平对于人类健康具有重要意义。为更好地了解影响植物富硒效果的因素,综述了土壤中硒的赋存形态,重点阐述了植物对土壤不同形态硒的吸收、迁移和代谢过程,并由此对硒生物强化未来的研究方向进行了展望。
Selenate redistribution during aging in different Chinese soils and the dominant influential factors
DOI:S0045-6535(17)30709-9
PMID:28500973
[本文引用: 1]
To date, few works have attempted to determine the effect of soil types on Selenium aging process and the possible influential factors. In this study, the differences in Se speciation distribution and availability in 15 Chinese typical agricultural soils were investigated using spiked selenate for the entire year. Results evidenced that after one year of incubation, Se transformed from soluble fraction to Fe/Mn oxides and organic matter bound fractions in neutral or alkaline soils (pH 7.09-8.51) and from exchangeable fraction to residual fraction in acidic soils (pH 4.89-6.82). The available Se content in all soils declined rapidly at the initial stage of aging, with most of the neutral or alkaline soils reaching equilibrium after 109 d, whereas the acidic soils reached equilibrium after only 33-56 d. The available Se content in soil decreased constantly during the entire aging process in S4 (Xinjiang Gray desert soil), S12 (Anhui Yellow brown earths), and S15 (Hunan Krasnozems). Elovich model was the best model (R > 0.80) in describing the Se aging process. Estimated time for exogenous Se reaching the distribution of available Se in corresponding native soils extended from 9.7 y to 50.2 y, indicating a much longer time was required for spiked soil to reach equilibrium. Soil pH was the most significant factor directly and negatively influencing the aging process (p < 0.05), while organic matter played a dual role on Se speciation. Results could provide reference for the selection of unified equilibrium time on Se-spiked experiment.Copyright © 2017 Elsevier Ltd. All rights reserved.
Unequal functional redundancy between the two Arabidopsis thaliana high-affinity sulphate transporters SULTR1;1 and SULTR1;2
DOI:10.1111/j.1469-8137.2008.02604.x
PMID:18761637
[本文引用: 1]
* In Arabidopsis, SULTR1;1 and SULTR1;2 are two genes proposed to be involved in high-affinity sulphate uptake from the soil solution. We address here the specific issue of their functional redundancy for the uptake of sulphate and for the accumulation of its toxic analogue selenate with regard to plant growth and selenate tolerance. * Using the complete set of genotypes, including the wild-type, each one of the single sultr1;1 and sultr1;2 mutants and the resulting double sultr1;1-sultr1;2 mutant, we performed a detailed phenotypic analysis of root length, shoot biomass, sulphate uptake, sulphate and selenate accumulation and selenate tolerance. * The results all ordered the four different genotypes according to the same functional hierarchy. Wild-type and sultr1;1 mutant plants displayed similar phenotypes. By contrast, sultr1;1-sultr1;2 double-mutant plants showed the most extreme phenotype and the sultr1;2 mutant displayed intermediate performances. Additionally, the degree of selenate tolerance was directly related to the seedling selenate content according to a single sigmoid regression curve common to all the genotypes. * The SULTR1;1 and SULTR1;2 genes display unequal functional redundancy, which leaves open for SULTR1;1 the possibility of displaying an additional function besides its role in sulphate membrane transport.
Differential regulation of the expression of two high-affinity sulfate transporters,SULTR1.1 and SULTR1.2,in Arabidopsis
DOI:10.1104/pp.108.118612
PMID:18400935
[本文引用: 1]
The molecular mechanisms regulating the initial uptake of inorganic sulfate in plants are still largely unknown. The current model for the regulation of sulfate uptake and assimilation attributes positive and negative regulatory roles to O-acetyl-serine (O-acetyl-Ser) and glutathione, respectively. This model seems to suffer from exceptions and it has not yet been clearly validated whether intracellular O-acetyl-Ser and glutathione levels have impacts on regulation. The transcript level of the two high-affinity sulfate transporters SULTR1.1 and SULTR1.2 responsible for sulfate uptake from the soil solution was compared to the intracellular contents of O-acetyl-Ser, glutathione, and sulfate in roots of plants submitted to a wide diversity of experimental conditions. SULTR1.1 and SULTR1.2 were differentially expressed and neither of the genes was regulated in accordance with the current model. The SULTR1.1 transcript level was mainly altered in response to the sulfur-related treatments. Split-root experiments show that the expression of SULTR1.1 is locally regulated in response to sulfate starvation. In contrast, accumulation of SULTR1.2 transcripts appeared to be mainly related to metabolic demand and is controlled by photoperiod. On the basis of the new molecular insights provided in this study, we suggest that the expression of the two transporters depends on different regulatory networks. We hypothesize that interplay between SULTR1.1 and SULTR1.2 transporters could be an important mechanism to regulate sulfate content in the roots.
Characterization of a selenate-resistant Arabidopsis mutant:root growth as a potential target for selenate toxicity
DOI:10.1104/pp.106.091462 URL [本文引用: 1]
Phloem-localizing sulfate transporter,Sultr1;3,mediates re-distribution of sulfur from source to sink organs in Arabidopsis
DOI:10.1104/pp.014712
PMID:12692311
[本文引用: 1]
For the effective recycling of nutrients, vascular plants transport pooled inorganic ions and metabolites through the sieve tube. A novel sulfate transporter gene, Sultr1;3, was identified as an essential member contributing to this process for redistribution of sulfur source in Arabidopsis. Sultr1;3 belonged to the family of high-affinity sulfate transporters, and was able to complement the yeast sulfate transporter mutant. The fusion protein of Sultr1;3 and green fluorescent protein was expressed by the Sultr1;3 promoter in transgenic plants, which revealed phloem-specific expression of Sultr1;3 in Arabidopsis. Sultr1;3-green fluorescent protein was found in the sieve element-companion cell complexes of the phloem in cotyledons and roots. Limitation of external sulfate caused accumulation of Sultr1;3 mRNA both in leaves and roots. Movement of (35)S-labeled sulfate from cotyledons to the sink organs was restricted in the T-DNA insertion mutant of Sultr1;3. These results provide evidence that Sultr1;3 transporter plays an important role in loading of sulfate to the sieve tube, initiating the source-to-sink translocation of sulfur nutrient in Arabidopsis.
Selenium promotes sulfur accumulation and plant growth in wheat (Triticum aestivum)
DOI:10.1111/ppl.2016.158.issue-1 URL [本文引用: 1]
Influence of sulfate supply on selenium uptake dynamics and expression of sulfate/selenate transporters in selenium hyperaccumulator and nonhyperaccumulator Brassicaceae
DOI:10.1111/nph.2018.217.issue-1 URL [本文引用: 1]
SULTR3; 1 is a chloroplast- localized sulfate transporter in Arabidopsis thaliana
DOI:10.1111/tpj.2013.73.issue-4 URL [本文引用: 1]
Root-to-shoot transport of sulfate in Arabidopsis. Evidence for the role of SULTR3;5 as a component of low-affinity sulfate transport system in the root vasculature
DOI:10.1104/pp.104.045625
PMID:15531709
[本文引用: 1]
Xylem transport of sulfate regulates distribution of sulfur in vascular plants. Here, we describe SULTR3;5 as an essential component of the sulfate transport system that facilitates the root-to-shoot transport of sulfate in the vasculature. In Arabidopsis (Arabidopsis thaliana), SULTR3;5 was colocalized with the SULTR2;1 low-affinity sulfate transporter in xylem parenchyma and pericycle cells in roots. In a yeast (Saccharomyces cerevisiae) expression system, sulfate uptake was hardly detectable with SULTR3;5 expression alone; however, cells coexpressing both SULTR3;5 and SULTR2;1 showed substantial uptake activity that was considerably higher than with SULTR2;1 expression alone. The V(max) value of sulfate uptake activity with SULTR3;5-SULTR2;1 coexpression was approximately 3 times higher than with SULTR2;1 alone. In Arabidopsis, the root-to-shoot transport of sulfate was restricted in the sultr3;5 mutants, under conditions of high SULTR2;1 expression in the roots after sulfur limitation. These results suggested that SULTR3;5 is constitutively expressed in the root vasculature, but its function to reinforce the capacity of the SULTR2;1 low-affinity transporter is only essential when SULTR2;1 mRNA is induced by sulfur limitation. Consequently, coexpression of SULTR3;5 and SULTR2;1 provides maximum capacity of sulfate transport activity, which facilitates retrieval of apoplastic sulfate to the xylem parenchyma cells in the vasculature of Arabidopsis roots and may contribute to the root-to-shoot transport of sulfate.
Selenium promotes sulfur accumulation and plant growth in wheat (Triticum aestivum)
DOI:10.1111/ppl.2016.158.issue-1 URL [本文引用: 1]
Vacuolar sulfate transporters are essential determinants controlling internal distribution of sulfate in Arabidopsis
DOI:10.1105/tpc.104.023960
PMID:15367713
[本文引用: 1]
Uptake of external sulfate from the environment and use of internal vacuolar sulfate pools are two important aspects of the acquisition of sulfur for metabolism. In this study, we demonstrated that the vacuolar SULTR4-type sulfate transporter facilitates the efflux of sulfate from the vacuoles and plays critical roles in optimizing the internal distribution of sulfate in Arabidopsis thaliana. SULTR4;1-green fluorescent protein (GFP) and SULTR4;2-GFP fusion proteins were expressed under the control of their own promoters in transgenic Arabidopsis. The fusion proteins were accumulated specifically in the tonoplast membranes and were localized predominantly in the pericycle and xylem parenchyma cells of roots and hypocotyls. In roots, SULTR4;1 was constantly accumulated regardless of the changes of sulfur conditions, whereas SULTR4;2 became abundant by sulfur limitation. In shoots, both transporters were accumulated by sulfur limitation. Vacuoles isolated from callus of the sultr4;1 sultr4;2 double knockout showed excess accumulation of sulfate, which was substantially decreased by overexpression of SULTR4;1-GFP. In seedlings, the supplied [(35)S]sulfate was retained in the root tissue of the sultr4;1 sultr4;2 double knockout mutant. Comparison of the double and single knockouts suggested that SULTR4;1 plays a major role and SULTR4;2 has a supplementary function. Overexpression of SULTR4;1-GFP significantly decreased accumulation of [(35)S]sulfate in the root tissue, complementing the phenotype of the double mutant. These results suggested that SULTR4-type transporters, particularly SULTR4;1, actively mediate the efflux of sulfate from the vacuole lumen into the cytoplasm and influence the capacity for vacuolar storage of sulfate in the root tissue. The efflux function will promote rapid turnover of sulfate from the vacuoles particularly in the vasculature under conditions of low-sulfur supply, which will optimize the symplastic (cytoplasmic) flux of sulfate channeled toward the xylem vessels.
Investigation of selenium tolerance mechanisms in Arabidopsis thaliana
DOI:10.1111/ppl.2006.128.issue-2 URL [本文引用: 1]
Effects of selenate on Se,flavonoid,and glucosinolate in broccoli florets by combined transcriptome and metabolome analyses
DOI:10.1016/j.foodres.2021.110463 URL [本文引用: 1]
Transport processes of solutes across the vacuolar membrane of higher plants
DOI:10.1093/pcp/pcd059 URL [本文引用: 1]
Transport of selenate and selenite into astragalus roots
DOI:10.1104/pp.44.6.893 URL [本文引用: 1]
Selenium uptake,translocation and speciation in wheat supplied with selenate or selenite
DOI:10.1111/nph.2008.178.issue-1 URL [本文引用: 3]
Overexpression of the phosphate transporter gene OsPT8 improves the Pi and selenium contents in Nicotiana tabacum
DOI:10.1016/j.envexpbot.2017.02.011 URL [本文引用: 1]
OsPT2,a phosphate transporter,is involved in the active uptake of selenite in rice
DOI:10.1111/nph.2014.201.issue-4 URL [本文引用: 1]
Involvement of silicon influx transporter OsNIP2; 1 in selenite uptake in rice
DOI:10.1104/pp.110.157867
URL
[本文引用: 1]
Rice (Oryza sativa) as a staple food, provides a major source of dietary selenium (Se) for humans, which essentially requires Se, however, the molecular mechanism for Se uptake is still poorly understood. Herein, we show evidence that the uptake of selenite, a main bioavailable form of Se in paddy soils, is mediated by a silicon (Si) influx transporter Lsi1 (OsNIP2;1) in rice. Defect of OsNIP2;1 resulted in a significant decrease in the Se concentration of the shoots and xylem sap when selenite was given. However, there was no difference in the Se concentration between the wild-type rice and mutant of OsNIP2;1 when selenate was supplied. A short-term uptake experiment showed that selenite uptake greatly increased with decreasing pH in the external solution. Si as silicic acid did not inhibit the Se uptake from selenite in both rice and yeast (Saccharomyces cerevisiae) at low pHs. Expression of OsNIP2;1 in yeast enhanced the selenite uptake at pH 3.5 and 5.5 but not at pH 7.5. On the other hand, defect of Si efflux transporter Lsi2 did not affect the uptake of Se either from selenite or selenate. Taken together, our results indicate that Si influx transporter OsNIP2;1 is permeable to selenite.
Effects of selenium fertilizer on grain yield,Se uptake and distribution in common buckwheat (Fagopyrum esculentum Moench)
DOI:10.17221/284/2015-PSE URL [本文引用: 1]
Do selenium hyperaccumulators affect selenium speciation in neighboring plants and soil? An X-ray microprobe analysis
DOI:10.1080/15226514.2014.987374
PMID:26030363
[本文引用: 1]
Neighbors of Se hyperaccumulators Stanleya pinnata and Astragalus bisulcatus were found earlier to have elevated Se levels. Here we investigate whether Se hyperaccumulators affect Se localization and speciation in surrounding soil and neighboring plants. X-ray fluorescence mapping and X-ray absorption near-edge structure spectroscopy were used to analyze Se localization and speciation in leaves of Artemisia ludoviciana, Symphyotrichum ericoides and Chenopodium album growing next to Se hyperaccumulators or non-accumulators at a seleniferous site. Regardless of neighbors, A. ludoviciana, S. ericoides and C. album accumulated predominantly (73-92%) reduced selenocompounds with XANES spectra similar to the C-Se-C compounds selenomethionine and methyl-selenocysteine. Preliminary data indicate that the largest Se fraction (65-75%), both in soil next to hyperaccumulator S. pinnata and next to nonaccumulator species was reduced Se with spectra similar to C-Se-C standards. These same C-Se-C forms are found in hyperaccumulators. Thus, hyperaccumulator litter may be a source of organic soil Se, but soil microorganisms may also contribute. These findings are relevant for phytoremediation and biofortification since organic Se is more readily accumulated by plants, and more effective for dietary Se supplementation.
水稻对有机态硒的吸收与积累
DOI:10.11924/j.issn.1000-6850.casb16050144
[本文引用: 1]
采用盆栽试验,研究叶面喷施有机态硒(Se-Met)对水稻吸收、积累硒的影响,明确以酵母硒作为有机硒源对提高植物富硒能力的影响。试验结果表明:叶面施用有机硒后,植株不同部位吸收、积累硒能力不同,按照叶、茎、穗顺序逐渐降低;植株中硒积累量以孕穗期施用高于苗期喷硒;籽粒中硒积累量以抽穗期施用最高,苗期喷硒最低;水稻籽粒中硒含量以稻壳中最高,是大米中硒含量的1.1-4倍左右;不同用喷施量表明,有机硒用量8 g.亩<sub>-1</sub>,更有利于籽粒中硒的积累。有机硒作为一种外源硒,可以满足水稻生长对硒营养的需求,提高籽粒中硒的富集。
Plant uptake and translocation of inorganic and organic forms of selenium
DOI:10.1007/s00244-013-9926-0
PMID:23793939
[本文引用: 1]
Selenium (Se) plays a role in human health: It is an essential trace element but can be toxic if too much is consumed. The aim of this study was to determine which species of Se are most rapidly taken up and translocated to above-ground plant tissues. Specifically, we wished to determine if organic forms of Se in an exposure solution can contribute to the amount of Se found in shoot tissue. Durum wheat (Triticum turgidum) and spring canola (Brassica napus) were grown hydroponically, and young seedlings were exposed to 0.5 or 5.0 μM Se as selenate, selenite, seleno-methionione, or seleno-cystine for ≤300 min. Canola accumulated more Se than wheat, although the difference depended on Se speciation of the exposure solution. Organic forms of Se were taken up at a greater rate than inorganic forms. When exposed to 5.0 μM Se, the rate of uptake of selenite was 1.5- (canola) or 5-fold (wheat) greater than the rate of uptake of selenate, whereas seleno-methionine was taken up 40- (canola) or 100-fold (wheat) faster and seleno-cystine 2- (wheat) to 20-fold (canola) faster. Plants exposed to seleno-methionine had the highest shoot concentrations of Se even though selenate was more mobile once taken up; in plants exposed to selenate >50% of accumulated Se was translocated to shoot tissue. Because organic forms of Se (especially seleno-methionine) can be readily taken up and translocated to above-ground tissues of wheat and canola, these Se species should be considered when attempting to predict Se accumulation in above-ground plant tissues.
Organic selenium distribution in selected California soils
DOI:10.2136/sssaj1990.03615995005400040007x URL [本文引用: 1]
NRT1.1B improves selenium concentrations in rice grains by facilitating selenomethinone translocation
DOI:10.1111/pbi.13037
PMID:30466149
[本文引用: 1]
Selenium (Se) is an essential trace element for humans and other animals, yet approximately one billion people worldwide suffer from Se deficiency. Rice is a staple food for over half of the world's population that is a major dietary source of Se. In paddy soils, rice roots mainly take up selenite. Se speciation analysis indicated that most of the selenite absorbed by rice is predominantly transformed into selenomethinone (SeMet) and retained in roots. However, the mechanism by which SeMet is transported in plants remains largely unknown. In this study, SeMet uptake was found to be an energy-dependent symport process involving H transport, with neutral amino acids strongly inhibiting SeMet uptake. We further revealed that NRT1.1B, a member of rice peptide transporter (PTR) family which plays an important role in nitrate uptake and transport in rice, displays SeMet transport activity in yeast and Xenopus oocyte. The uptake rate of SeMet in the roots and its accumulation rate in the shoots of nrt1.1b mutant were significantly repressed. Conversely, the overexpression of NRT1.1B in rice significantly promoted SeMet translocation from roots to shoots, resulting in increased Se concentrations in shoots and rice grains. With vascular-specific expression of NRT1.1B, the grain Se concentration was 1.83-fold higher than that of wild type. These results strongly demonstrate that NRT1.1B holds great potential for the improvement of Se concentrations in grains by facilitating SeMet translocation, and the findings provide novel insight into breeding of Se-enriched rice varieties.© 2018 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Foliar water and solute absorption:an update
DOI:10.1111/tpj.v105.4 URL [本文引用: 1]
Selection pressures on stomatal evolution
DOI:10.1046/j.0028-646X.2001.00334.x
PMID:33863217
[本文引用: 1]
Fossil evidence shows that stomata have occurred in sporophytes and (briefly) gametophytes of embryophytes during the last 400 m yr. Cladistic analyses with hornworts basal are consistent with a unique origin of stomata, although cladograms with hornworts as the deepest branching embryophytes require loss of stomata early in the evolution of liverworts. Functional considerations suggest that stomata evolved from pores in the epidermis of plant organs which were at least three cell layers thick and had intercellular gas spaces and a cuticle; an endohydric conducting system would not have been necessary for low-growing rhizophytes, especially in early Palaeozoic CO -rich atmospheres. The 'prestomatal state' (pores) would have permitted higher photosynthetic rates per unit ground area. Functional stomata, and endohydry, permit the evolution of homoiohydry and the loss of vegetative desiccation tolerance and plants > 1 m tall. Stomatal functioning would then have involved maintenance of hydration, and restricting the occurrence of xylem embolism, under relatively desiccating conditions at the expense of limiting carbon acquisition. The time scale of environmental fluctuations over which stomatal responses can maximize carbon gain per unit water loss varies among taxa and life forms. Contents Summary 371 I. Introduction 371 II. Monophyly of stomata? 372 III. Roles of stomata in extant plants 373 IV. Ecophysiology of ancestrally astomatous terrestrial plants 375 V. Evolution of stomata 379 VI. Ecophysiological implications of losses of stomata 382 VII. Conclusions 384 Acknowledgements 384 References 384.
Size exclusion limits and lateral heterogeneity of the stomatal foliar uptake pathway for aqueous solutes and water-suspended nanoparticles
SULTR3s function in chloroplast sulfate uptake and affect ABA biosynthesis and the stress response
DOI:10.1104/pp.18.01439
PMID:30837346
[本文引用: 1]
Plants are major sulfur reducers in the global sulfur cycle. Sulfate, the major natural sulfur source in soil, is absorbed by plant roots and transported into plastids, where it is reduced and assimilated into Cys for further metabolic processes. Despite its importance, how sulfate is transported into plastids is poorly understood. We previously demonstrated using single Arabidopsis () genetic mutants that each member of the sulfate transporter (SULTR) subfamily 3 was able to transport sulfate across the chloroplast envelope membrane. To resolve the function of SULTR3s, we constructed a quintuple mutant completely knocking out all five members of the subfamily. Here we report that all members of the subfamily show chloroplast membrane localization. Sulfate uptake by chloroplasts of the quintuple mutant is reduced by more than 50% compared with the wild type. Consequently, Cys and abscisic acid (ABA) content are reduced to ∼67 and ∼20% of the wild-type level, respectively, and strong positive correlations are found among sulfate, Cys, and ABA content. The quintuple mutant shows obvious growth retardation with smaller rosettes and shorter roots. Seed germination of the quintuple mutant is hypersensitive to exogenous ABA and salt stress, but is rescued by sulfide supplementation. Furthermore, sulfate-induced stomatal closure is abolished in the quintuple mutant, strongly suggesting that chloroplast sulfate is required for stomatal closure. Our genetic analyses unequivocally demonstrate that sulfate transporter subfamily 3 is responsible for more than half of the chloroplast sulfate uptake and influences downstream sulfate assimilation and ABA biosynthesis.© 2019 American Society of Plant Biologists. All Rights Reserved.
Selenium uptake,translocation,assimilation and metabolic fate in plants
The chemical and physical resemblance between selenium (Se) and sulfur (S) establishes that both these elements share common metabolic pathways in plants. The presence of isologous Se and S compounds indicates that these elements compete in biochemical processes that affect uptake, translocation and assimilation throughout plant development. Yet, minor but crucial differences in reactivity and other metabolic interactions infer that some biochemical processes involving Se may be excluded from those relating to S. This review examines the current understanding of physiological and biochemical relationships between S and Se metabolism by highlighting their similarities and differences in relation to uptake, transport and assimilation pathways as observed in Se hyperaccumulator and non-accumulator plant species. The exploitation of genetic resources used in bioengineering strategies of plants is illuminating the function of sulfate transporters and key enzymes of the S assimilatory pathway in relation to Se accumulation and final metabolic fate. These strategies are providing the basic framework by which to resolve questions relating to the essentiality of Se in plants and the mechanisms utilized by Se hyperaccumulators to circumvent toxicity. In addition, such approaches may assist in the future application of genetically engineered Se accumulating plants for environmental renewal and human health objectives.
Sulfate reduction in higher plants:molecular evidence for a novel 5′-adenylylsulfate reductase
Analysis of sulfur and selenium assimilation in Astragalus plants with varying capacities to accumulate selenium
DOI:10.1111/tpj.2005.42.issue-6 URL [本文引用: 1]
Superoxide generated from the glutathione-mediated reduction of selenite damages the iron-sulfur cluster of chloroplastic ferredoxin
DOI:10.1016/j.plaphy.2016.05.004 URL [本文引用: 1]
Acid-volatile selenium formation catalyzed by glutathione reductase
The production of acid-volatile selenide (apparently H2Se) was catalyzed by glutathione reductase in an anaerobic system containing 20 mM glutathione, 0.05 mM sodium selenite, a TPNH-generating system, and microgram quantities of highly purified yeast glutathione reductase. H2Se production in this system was proportional to glutathione reductase concentration and was maximal at pH 7. Significant nonenzymic H2Se production occurred in the system lacking glutathione reductase and TNPH. A concentration of arsenite (0.1 mM) which does not inhibit glutathione reductase inhibited selenide volatilization, as did bovine serum albumin (1.67 mg/ml). Both appear to inhibit Se volatilization by reacting with the selenide product(s). The selenotrisulfide derivative of glutathione (GSSeSG) was readily converted to H2Se by glutathione reductase and TPNH without the addition of glutathione. These results suggest that GSSeSG formed nonenzymically from glutathione and selenic undergoes stepwise reduction by glutathione reductase (or excess GSH) to GSSeH and finally to H2Se. The same pathway operates when glutathione is used as the reducing agent but to a lesser extent.
Cysteine synthesis in plants:protein- protein interactions of serine acetyltransferase from Arabidopsis thaliana
DOI:10.1046/j.1365-313X.1997.11020251.x URL [本文引用: 1]
Incorporation of cysteine and selenocysteine into cystathionine and selenocystathionine by crude extracts of spinach
DOI:10.1016/0031-9422(88)80747-7 URL [本文引用: 1]
L-Selenohomocysteine:one-step synthesis from L-selenomethionine and kinetic analysis as substrate for methionine synthases
A single-step convenient synthesis of L-selenohomocysteine (SeHcy) from L-selenomethionine (SeMet) using sodium in liquid ammonia is described. Methionine synthases convert SeHcy to SeMet at rates comparable to their rates of conversion of L-homocysteine (Hcy) to L-methionine (Met). This study suggests that SeHcy generated from SeMet metabolism can be efficiently recycled to SeMet in mammals.
An essential role of S-adenosyl- L-methionine:L-methionine S-methyltransferase in selenium volatilization by plants. Methylation of selenomethionine to selenium-methyl-L-selenium-methionine,the precursor of volatile selenium
DOI:10.1104/pp.001693
URL
[本文引用: 1]
Selenium (Se) phytovolatilization, the process by which plants metabolize various inorganic or organic species of Se (e.g. selenate, selenite, and Se-methionine [Met]) into gaseous Se forms (e.g. dimethylselenide), is a potentially important means of removing Se from contaminated environments. Before attempting to genetically enhance the efficiency of Se phytovolatilization, it is essential to elucidate the enzymatic pathway involved and to identify its rate-limiting steps. The present research tested the hypothesis thatS-adenosyl-l-Met:l-MetS-methyltransferase (MMT) is the enzyme responsible for the methylation of Se-Met to Se-methyl Se-Met (SeMM). To this end, we identified and characterized an Arabidopsis T-DNA mutant knockout for MMT. The lack of MMT in the Arabidopsis T-DNA mutant plant resulted in an almost complete loss in its capacity for Se volatilization. Using chemical complementation with SeMM, the presumed enzymatic product of MMT, we restored the capacity of the MMT mutant to produce volatile Se. Overexpressing MMT from Arabidopsis in Escherichia coli, which is not known to have MMT activity, produced up to 10 times more volatile Se than the untransformed strain when both were supplied with Se-Met. Thus, our results provide in vivo evidence that MMT is the key enzyme catalyzing the methylation of Se-Met to SeMM.
Volatile selenium in higher plants the production of dimethyl selenide in cabbage leaves by enzymatic cleavage of Se-methyl selenomethionine selenonium salt
DOI:10.1007/BF00011413 URL [本文引用: 1]
Selenium assimilation and volatilization from dimethylselenoniopropionate by Indian Mustard
Earlier work from our laboratory on Indian mustard (Brassica juncea L.) identified the following rate-limiting steps for the assimilation and volatilization of selenate to dimethyl selenide (DMSe): (a) uptake of selenate, (b) activation of selenate by ATP sulfurylase, and (b) conversion of selenomethionine (SeMet) to DMSe. The present study showed that shoots of selenate-treated plants accumulated very low concentrations of dimethylselenoniopropionate (DMSeP). Selenonium compounds such as DMSeP are the most likely precursors of DMSe. DMSeP-supplied plants volatilized Se at a rate 113 times higher than that measured from plants supplied with selenate, 38 times higher than from selenite, and six times higher than from SeMet. The conversion of SeMet to selenonium compounds such as DMSeP is likely to be rate-limiting for DMSe production, but not the formation of DMSe from DMSeP because DMSeP was the rate of Se volatilization from faster than from SeMet and SeMet (but no DMSeP) accumulated in selenite- or SeMet-supplied wild-type plants and in selenate-supplied ATP-sulfurylase transgenic plants. DMSeP-supplied plants absorbed the most Se from the external medium compared with plants supplied with SeMet, selenate, or selenite; they also accumulated more Se in shoots than in roots as an unknown organic compound resembling a mixture of DMSeP and selenocysteine.
Overexpression of selenocysteine methyltransferase in Arabidopsis and Indian mustard increases selenium tolerance and accumulation
A major goal of phytoremediation is to transform fast-growing plants with genes from plant species that hyperaccumulate toxic trace elements. We overexpressed the gene encoding selenocysteine methyltransferase (SMT) from the selenium (Se) hyperaccumulator Astragalus bisulcatus in Arabidopsis and Indian mustard (Brassica juncea). SMT detoxifies selenocysteine by methylating it to methylselenocysteine, a nonprotein amino acid, thereby diminishing the toxic misincorporation of Se into protein. Our Indian mustard transgenic plants accumulated more Se in the form of methylselenocysteine than the wild type. SMT transgenic seedlings tolerated Se, particularly selenite, significantly better than the wild type, producing 3- to 7-fold greater biomass and 3-fold longer root lengths. Moreover, SMT plants had significantly increased Se accumulation and volatilization. This is the first study, to our knowledge, in which a fast-growing plant was genetically engineered to overexpress a gene from a hyperaccumulator in order to increase phytoremediation potential.
Overexpression of AtCpNifS enhances selenium tolerance and accumulation in Arabidopsis
DOI:10.1104/pp.105.068684
URL
[本文引用: 1]
Selenium (Se) is an essential element for many organisms but is toxic at higher levels. CpNifS is a chloroplastic NifS-like protein in Arabidopsis (Arabidopsis thaliana) that can catalyze the conversion of cysteine into alanine and elemental sulfur (S0) and of selenocysteine into alanine and elemental Se (Se0). We overexpressed CpNifS to investigate the effects on Se metabolism in plants. CpNifS overexpression significantly enhanced selenate tolerance (1.9-fold) and Se accumulation (2.2-fold). CpNifS overexpressors showed significantly reduced Se incorporation into protein, which may explain their higher Se tolerance. Also, sulfur accumulation was enhanced by approximately 30% in CpNifS overexpressors, both on media with and without selenate. Root transcriptome changes in response to selenate mimicked the effects observed under sulfur starvation. There were only a few transcriptome differences between CpNifS-overexpressing plants and wild type, besides the 25- to 40-fold increase in CpNifS levels. Judged from x-ray analysis of near edge spectrum, both CpNifS overexpressors and wild type accumulated mostly selenate (SeVI). In conclusion, overexpression of this plant NifS-like protein had a pronounced effect on plant Se metabolism. The observed enhanced Se accumulation and tolerance of CpNifS overexpressors show promise for use in phytoremediation.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}