


工业大麻(Cannabis sativa L.)是一种生长快速、根系庞大且生物量高的植物,对重金属耐受性好,被应用于修复土壤重金属污染[10]。大麻二酚(cannabidiol,CBD)是工业大麻的主要活性成分,具有抗炎、抗氧化、抗凋亡和抗癌等活性作用,在医疗[11](神经系统、肿瘤等疾病)领域和护肤品行业[12]有着巨大的应用潜力。CBD在机体内能够通过调节抗氧化酶活性、抗氧化物质含量[13-14]和细胞内锌离子浓度维持氧化平衡;或利用自身的强还原性直接清除ROS等方式应对氧化应激损伤,但CBD对植物生长发育的作用及提高植物重金属抗性的研究鲜见报道。为此,本文以烟草为研究对象,探讨外源喷施CBD对Cd和Cr胁迫下烟株生长和抗氧化系统的影响,以期为CBD的应用及植物重金属防控提供依据和参考。
1 材料与方法
1.1 试验材料
供试烟草品种为K326(玉溪中烟种子有限责任公司)。二水乙酸镉由天津市科密欧化学试剂有限公司提供,重铬酸钾由西陇化工股份有限公司提供。CBD购买于云南汉盟制药有限公司,纯度为99%。试验土壤为红壤土,采自云南自然背景低肥力土壤,装盆前过2mm筛,充分混匀。土壤pH 6.84、有机质6.97g/kg、有效磷11.56mg/kg、速效钾101.33mg/kg、碱解氮18.91mg/kg、全铬(Cr)84.24mg/kg、全镉(Cd)0.27mg/kg,Cr和Cd均低于GB 15618-2018《土壤环境质量农用地土壤污染风险管控标准(试行)》规定的土壤污染风险筛选值(Cr 200mg/kg,Cd 0.3mg/kg,6.5<pH≤7.5)。
1.2 试验设计
盆栽试验于2021年5-7月在云南农业大学烟草学院设施大棚内进行。Cd和Cr土壤添加浓度分别设置为60和90mg/kg,CBD浓度为0.0、1.0、5.0和25.0mg/L,无Cd、Cr胁迫且无CBD处理为对照(CK),共9个处理,每个处理重复5盆。Cd、Cr分别用适量二水乙酸镉和重铬酸钾配制成1L水溶液,于移栽前2个月以模拟污灌的方式一次性施入装有10kg风干过筛土的花盆(上下口径、高度为34cm×19cm×23cm,下置接水盘)中,之后每天浇适量水以保持土壤湿润,并于移栽前混匀。将漂浮育苗培育的烟草壮苗(苗龄55d)移栽入花盆,环施45g烟草专用复合肥[m(N): m(P2O5):m(K2O)=15:15:18]作基肥。后期正常管理,视土壤干湿情况定期浇水、除草。移栽后50d,于上午第1次喷施CBD于叶片正反面,以湿润不流滴为宜,3d后喷施第2次。间隔3d后,用打孔器均匀地取一定质量的中部叶(自上而下7、8位叶)保存到液氮中,用于生理指标测定,剩余部分继续生长30d后采收,测定生物量和重金属含量。CBD溶液的配制,采用含0.1%的二甲基亚砜和0.07%的吐温80混合水溶液溶解而成。
1.3 测定项目及方法
于移栽后80d,各处理选取3株分部位收获,先用自来水洗净根、茎、叶表面的土壤,然后用0.01mol/L EDTA与去离子水洗净植株,用滤纸吸干水分,于105℃下杀青30min,65℃烘干至恒重,称量每株烟地上、地下部分干质量。将干燥后样品混合均匀,采用电感耦合等离子体质谱法测定各部位中的Cd和Cr含量。
耐受系数=各处理烟株地上(下)部生物量/对照烟株地上(下)生物量;转运系数(translocation factor,TF)=烟株地上部分Cd或Cr含量/地下部分Cd或Cr含量;富集系数=植物地上(或地下)部Cd或Cr含量/土壤中Cd或Cr含量;单株富集量=单株生物量×单株Cd或Cr含量。
1.4 数据处理
采用Origin 2018和SPSS 25软件进行统计分析及差异显著性检验。对不同处理的数据进行单因素方差分析(ANOVA)和Duncan多重比较(P<0.05)。数据以平均值±标准差表示。采用Pearson法进行相关性分析。
2 结果与分析
2.1 CBD对Cd和Cr胁迫下烟株生物量的影响
由表1可知,Cd和Cr胁迫下,烟株生物量显著降低。但随CBD浓度增加,烟株干重和重金属耐性均先增加后降低。Cd胁迫下,1.0mg/LCBD处理烟株干重最大,分别比单一Cd处理显著提高13.93%(地上)和31.63%(地下)。Cr胁迫下,5mg/L CBD处理效果最佳,烟株干重和耐受系数分别比单一Cr处理显著提高21.97%和21.92%(地上)、26.03%和25.97%(地下)。而25.0mg/L CBD处理时,烟株干重和耐受系数均与单独Cd、Cr处理无显著差异。
表1 CBD对Cd和Cr胁迫下烟株生物量的影响
Table 1
| 重金属 Heavy metal (mg/kg) | CBD浓度 CBD concentration (mg/L) | 干重(g/株)Dry weight (g/plant) | 耐受系数Tolerance coefficient | |||
|---|---|---|---|---|---|---|
| 地上部分 Upper ground | 地下部分 Under ground | 地上部分 Upper ground | 地下部分 Under ground | |||
| CK | 51.47±0.82a | 7.62±0.38a | − | − | ||
| Cd (60) | 0.0 | 41.00±1.17c | 5.47±0.44c | 0.80±0.03b | 0.72±0.09b | |
| 1.0 | 46.71±1.02b | 7.20±0.46a | 0.91±0.02a | 0.94±0.06a | ||
| 5.0 | 42.81±1.18c | 7.04±0.62ab | 0.83±0.01b | 0.92±0.08a | ||
| 25.0 | 41.91±1.05c | 6.18±1.05bc | 0.81±0.03b | 0.81±0.10ab | ||
| Cr (90) | 0.0 | 37.68±0.68d | 5.84±0.24b | 0.73±0.00c | 0.77±0.01b | |
| 1.0 | 39.47±0.78c | 6.32±0.28b | 0.77±0.01b | 0.83±0.04b | ||
| 5.0 | 45.96±0.48b | 7.36±0.28a | 0.89±0.02a | 0.97±0.06a | ||
| 25.0 | 38.11±0.69d | 6.47±0.45b | 0.74±0.00c | 0.85±0.06b | ||
不同小写字母表示处理间差异显著(P < 0.05),下同
Different lowercase letters indicate significant difference between treatments (P < 0.05), the same below
2.2 CBD对Cd和Cr胁迫下烟株吸收Cd和Cr情况的影响
由表2可知,单独Cd和Cr胁迫下,烟株体内的Cd、Cr积累量大幅增加,分别是对照的19.73倍和1.85倍,且Cd含量地上部>地下部,而Cr含量则是地上部<地下部。烟株地上部、地下部Cd的富集系数分别为0.630~0.750、0.210~0.300,Cr分别为0.010~0.013、0.130~0.250。喷施适量的CBD能显著降低烟株中Cd和Cr积累量,但会提高Cd、Cr的转运系数。1.0mg/L CBD处理烟株地下部Cd含量最低,比单独Cd处理显著降低了30.00%,转运系数却显著增加了20.00%;而不同浓度CBD处理烟株的地上部Cd含量没有显著差异。1.0mg/L CBD处理地上部和5.0mg/L CBD处理地下部Cr含量最低,分别比单一胁迫降低了22.17%和47.72%。所有处理中Cr的转运系数均低于0.1,说明Cr主要富集在根部,只有微量转向地上部分。
表2 CBD对Cd和Cr胁迫下烟株吸收Cd、Cr情况的影响
Table 2
| 重金属 Heavy mental (mg/kg) | CBD浓度 CBD concentration (mg/L) | Cd或Cr含量 Content of Cd or Cr (mg/kg DW) | 富集系数 Bioaccumulation factor | Cd/Cr TF | Cd或Cr单株积累量 Cd or Cr accumulation per plant (mg) | |||
|---|---|---|---|---|---|---|---|---|
| 地上部分 Upper ground | 地下部分 Under ground | 地上部分 Upper ground | 地下部分 Under ground | |||||
| CK | 0.86±0.03c | 1.68±0.06d | − | − | 0.51±0.01c | 0.15±0.08d | ||
| Cd (60) | 0.0 | 45.41±1.39a | 18.18±0.30a | 0.750±0.02a | 0.300±0.00a | 2.50±0.11b | 2.96±0.14a | |
| 1.0 | 38.24±0.79b | 12.76±0.33c | 0.630±0.01b | 0.210±0.01c | 3.00±0.14a | 2.75±0.05b | ||
| 5.0 | 37.92±1.22b | 13.10±0.10c | 0.630±0.02b | 0.220±0.00c | 2.89±0.07a | 2.54±0.03c | ||
| 25.0 | 38.30±2.06b | 15.93±0.21b | 0.640±0.03b | 0.260±0.00b | 2.41±0.13b | 2.61±0.05c | ||
| CK | 1.40±0.06d | 16.49±0.31e | − | − | 0.08±0.00b | 1.06±0.00d | ||
| Cr (90) | 0.0 | 2.30±0.10a | 42.71±1.33a | 0.013±0.00a | 0.250±0.01a | 0.05±0.00d | 1.96±0.06a | |
| 1.0 | 1.79±0.05c | 26.76±0.41c | 0.010±0.00c | 0.150±0.00c | 0.07±0.00c | 1.31±0.04c | ||
| 5.0 | 2.08±0.11b | 22.33±0.76d | 0.012±0.00b | 0.130±0.00d | 0.09±0.01a | 1.30±0.04c | ||
| 25.0 | 2.15±0.09ab | 32.94±0.38b | 0.012±0.00b | 0.190±0.00b | 0.07±0.00c | 1.56±0.03b | ||
2.3 CBD对Cd和Cr胁迫下烟草叶片膜脂过氧化的影响
由图1可知,喷施中低浓度CBD(1.0和5.0mg/L)可显著降低Cd和Cr胁迫烟叶中活性氧和MDA含量。1.0mg/L CBD处理烟叶O2-·产生速率、H2O2和MDA含量分别比单独Cd显著降低31.75%、29.36%和14.40%。Cr胁迫下,5.0mg/L CBD处理O2-·产生速率和MDA含量最低,比单独Cr处理降低了29.21%和9.67%。而25.0mg/L CBD处理加重了叶片膜脂过氧化,其中MDA含量高于单独重金属处理。
图1
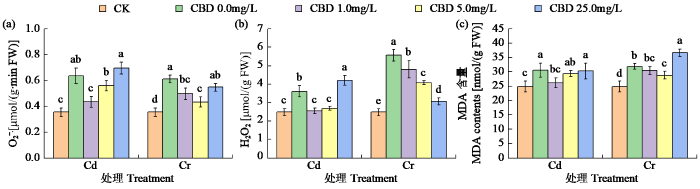
图1
CBD对Cd和Cr胁迫下烟叶中O2-·产生速率以及H2O2和MDA含量的影响
不同小写字母表示处理间差异显著(P < 0.05),下同
Fig.1
Effects of CBD on O2-· production rate, H2O2 and MDA contents in tobacco leaves under Cd and Cr stress
Different lowercase letters indicate significant difference (P < 0.05) between treatments, the same below
2.4 CBD对Cd和Cr胁迫下烟草叶片抗氧化酶活性的影响
由图2可知,Cd和Cr胁迫抑制了叶片中SOD、POD和PPO活性。随CBD浓度增加,叶片内抗氧化酶活性均先增后减。Cd胁迫下,1.0mg/L CBD处理使POD和PPO活性比单独Cd处理显著增加了10.59%和34.91%,5.0mg/L CBD处理使SOD活性提高了56.80%。Cr胁迫下,1.0mg/L CBD处理SOD和POD活性最高,分别比Cr处理显著提高17.40%和33.64%;5.0mg/L CBD处理的PPO活性显著提高29.60%。高浓度CBD(25.0mg/L)处理对Cd和Cr胁迫下烟草叶片酶活性的促进作用不显著,甚至还会产生抑制效果。
图2
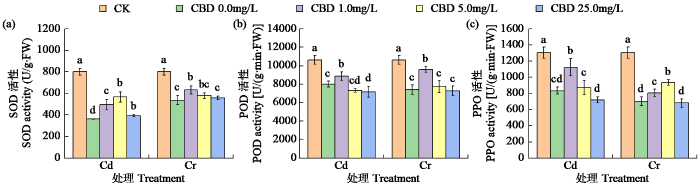
图2
CBD对Cd和Cr胁迫下烟叶中抗氧化酶活性的影响
Fig.2
Effects of CBD on antioxidant enzyme activities in tobacco leaves under Cd and Cr stress
2.5 CBD对Cd和Cr胁迫下烟叶中AsA-GSH循环的影响
2.5.1 对烟叶AsA和GSH含量、AsA/DHA和GSH/GSSG值的影响
由图3可知,Cd和Cr胁迫显著降低了叶片中AsA和GSH含量。喷施CBD可以促进2种抗氧化物质增加,但其促进效果随CBD浓度增加而减弱。Cd胁迫下,1.0mg/L CBD处理使AsA、GSH、AsA/DHA和GSH/GSSG分别比Cd处理显著提高了16.38%、103.17%、63.53%和467.07%。5.0mg/L CBD处理的叶片AsA/DHA和GSH/GSSG分别比单一Cr处理显著提高54.85%和100.19%。
图3
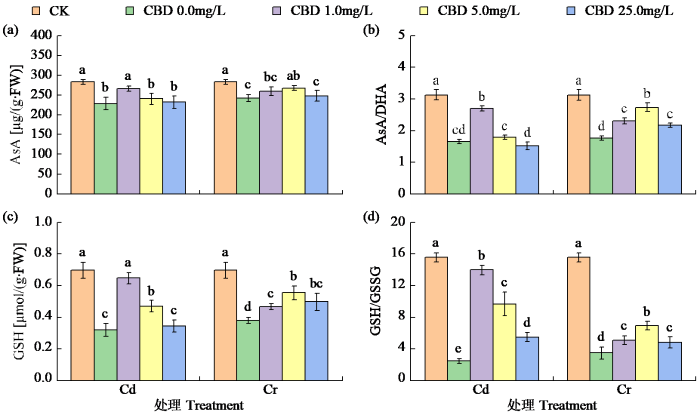
图3
CBD对Cd和Cr胁迫下烟叶AsA、AsA/DHA、GSH和GSH/GSSG的影响
Fig.3
Effects of CBD on AsA, AsA/DHA, GSH and GSH/GSSG in tobacco leaves under Cd and Cr stress
2.5.2 对烟叶APX和GR活性的影响
由图4可知,CBD对Cd和Cr胁迫烟叶的APX和GR活性存在低促高抑作用。Cd胁迫下,CBD浓度为1.0mg/L时,APX和GR活性最高,分别比单独Cd处理显著提高68.93%和25.8%,但CBD浓度大于5.0mg/L会对APX活性有显著的抑制作用。Cr胁迫下,CBD浓度为5.0mg/L时效果最佳,APX和GR活性分别比Cr处理显著提高35.58%和13.62%。
图4
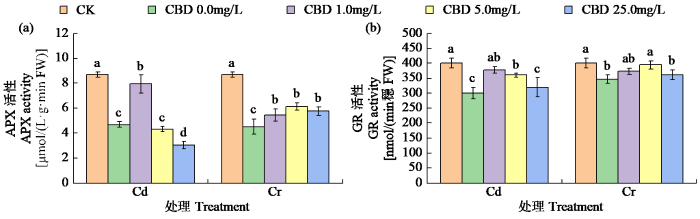
图4
CBD对Cd和Cr胁迫下烟叶APX和GR活性的影响
Fig.4
Effects of CBD on APX and GR activity of tobacco leaves under Cd and Cr stress
2.6 CBD浓度与烟叶氧化系统各指标和单株Cd、Cr积累量的相关性分析
由表3可知,CBD与烟株Cd、Cr积累量呈负相关。在Cd胁迫下,CBD与叶片POD、PPO活性呈显著负相关,与H2O2含量呈显著正相关,与O2-·产生速率呈极显著正相关。Cr胁迫下,除GSH和MDA外,CBD与其余指标皆呈负相关,其中与H2O2含量呈极显著负相关,与MDA含量呈极显著正相关。
表3 CBD浓度与烟叶氧化系统各指标和单株Cd、Cr积累量的相关系数
Table 3
| 重金属 Heavy metal | SOD | POD | PPO | AsA | GSH | H2O2 | O2-· | MDA | Cd或Cr积累量 Cd or Cr accumulation amount |
|---|---|---|---|---|---|---|---|---|---|
| Cd | -0.267 | -0.602* | -0.619* | -0.304 | -0.410 | 0.699* | 0.831** | 0.283 | -0.500 |
| Cr | -0.221 | -0.430 | -0.379 | -0.164 | 0.335 | -0.876** | -0.199 | 0.795 ** | -0.055 |
“**”表示极显著相关(P < 0.01),“*”表示显著相关(P < 0.05)
“**”shows significant correlation at the 0.01 level,“*”shows significant correlation at the 0.05 level
2.7 烟叶抗氧化指标与膜脂过氧化指标、单株Cd和Cr积累量的相关性分析
图5
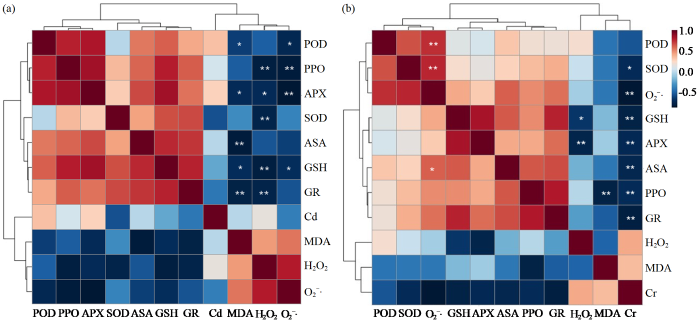
图5
Cd (a)、Cr (b)胁迫下烟叶抗氧化指标与膜脂过氧化指标、单株Cd和Cr积累量的相关性分析
“**”表示极显著相关(P < 0.01);“*”表示显著相关(P < 0.05)
Fig.5
Correlation analysis between antioxidant index and membrane lipid peroxidation index of tobacco leaves, Cd and Cr accumulation under Cd (a) and Cr (b) stress
“**”shows significant correlation at the 0.01 level;“*”shows significant correlation at the 0.05 level
3 讨论
植物主要通过根系吸收金属元素并转移到地上部。Cd在烟株中的移动性较好,主要积累在地上部,而Cr的移动性较低,故主要在根部积累。喷施CBD抑制了烟株对重金属的吸收,显著增加了烟株生物量。有学者[17]研究表明,油菜素内酯可以通过稳定植物细胞膜的电化学特性以及酶的活性来影响植物对重金属的吸收,故推测CBD可以发挥其稳定细胞膜的特性,以阻控烟株对Cd和Cr的吸收。另外,CBD可能与水杨酸、油菜素内酯调控植株Cd吸收类似[18-19],通过提高烟株的重金属耐受性,促进重金属胁迫下烟株对养分的吸收利用,改变烟株体内矿物质元素的分布,从而干扰Cd和Cr吸收相关的基因表达以及代谢通路,抑制植株对重金属的吸收。除此之外,植物激素水平[20]、根系分泌物[21]和土壤微生物群落结构[22]的改变,也会影响根系对重金属的吸收,但CBD是否会改变这些因素、其机理如何,有待进一步研究。
但CBD对Cd、Cr的缓解效应存在一定差异,对Cd的缓解效果优于Cr,且缓解Cd和Cr胁迫的最佳浓度有差异,这可能是因为Cd和Cr在烟株中的主要积累部位不同,导致二者叶片所受氧化损伤有差异;还有可能与烟株对不同类别重金属胁迫耐受性不同而引起的生理响应不同有关[33]。
4 结论
叶面喷施中低浓度的CBD(1.0、5.0mg/L)能激活SOD、POD、PPO、APX和GR活性,提高AsA和GSH含量,清除由Cd和Cr胁迫所产生的ROS,降低膜脂过氧化程度,抑制烟株对重金属的吸收,从而促进烟株生长。但高浓度CBD(25.0mg/L)对重金属胁迫的缓解作用下降,甚至会加剧损伤。由此可见,CBD在抗重金属胁迫方面具有潜在的应用前景,可以通过适当调节CBD的加入水平来缓解重金属对烟草生长发育的伤害。
参考文献
Heavy metal toxicity and sustainable interventions for their decontamination
DOI:10.1007/s42398-021-00164-y [本文引用: 1]
Methyl jasmonate alleviates arsenicinduced oxidative damage and modulates the ascorbate-glutathione cycle in oilseed rape roots
DOI:10.1007/s10725-017-0327-7 URL [本文引用: 1]
植物修复土壤重金属污染中外源物质的影响机制和应用研究进展
DOI:10.16258/j.cnki.1674-5906.2021.03.024
[本文引用: 1]

可下载PDF全文。
Bioaugmented phytoremediation of metal-contaminated soils and sediments by hemp and giant Reed
DOI:10.3389/fmicb.2021.645893
URL
[本文引用: 1]
We assessed the effects of EDTA and selected plant growth-promoting rhizobacteria (PGPR) on the phytoremediation of soils and sediments historically contaminated by Cr, Ni, and Cu. A total of 42 bacterial strains resistant to these heavy metals (HMs) were isolated and screened for PGP traits and metal bioaccumulation, and two Enterobacter spp. strains were finally selected. Phytoremediation pot experiments of 2 months duration were carried out with hemp (Cannabis sativa L.) and giant reed (Arundo donax L.) grown on soils and sediments respectively, comparing in both cases the effects of bioaugmentation with a single PGPR and EDTA addition on plant and root growth, plant HM uptake, HM leaching, as well as the changes that occurred in soil microbial communities (structure, biomass, and activity). Good removal percentages on a dry mass basis of Cr (0.4%), Ni (0.6%), and Cu (0.9%) were observed in giant reed while negligible values (&lt;100‰) in hemp. In giant reed, HMs accumulated differentially in plant (rhizomes &gt; &gt; roots &gt; leaves &gt; stems) with largest quantities in rhizomes (Cr 0.6, Ni 3.7, and Cu 2.2 g plant–1). EDTA increased Ni and Cu translocation to aerial parts in both crops, despite that in sediments high HM concentrations in leachates were measured. PGPR did not impact fine root diameter distribution of both crops compared with control while EDTA negatively affected root diameter class length (DCL) distribution. Under HM contamination, giant reed roots become shorter (from 5.2 to 2.3 mm cm–3) while hemp roots become shorter and thickened from 0.13 to 0.26 mm. A consistent indirect effect of HM levels on the soil microbiome (diversity and activity) mediated by plant response (root DCL distribution) was observed. Multivariate analysis of bacterial diversity and activity revealed not only significant effects of plant and soil type (rhizosphere vs. bulk) but also a clear and similar differentiation of communities between control, EDTA, and PGPR treatments. We propose root DCL distribution as a key plant trait to understand detrimental effect of HMs on microbial communities. Positive evidence of the soil-microbe-plant interactions occurring when bioaugmentation with PGPR is associated with deep-rooting perennial crops makes this combination preferable over the one with chelating agents. Such knowledge might help to yield better bioaugmented bioremediation results in contaminated sites.
In vitro effects of cannabidiol on activated immune-inflammatory pathways in major depressive patients and healthy controls
DOI:10.3390/ph15040405
URL
[本文引用: 1]
Major depressive disorder and major depressive episodes (MDD/MDE) are characterized by the activation of the immune–inflammatory response system (IRS) and the compensatory immune–regulatory system (CIRS). Cannabidiol (CBD) is a phytocannabinoid isolated from the cannabis plant, which is reported to have antidepressant-like and anti-inflammatory effects. The aim of the present study is to examine the effects of CBD on IRS, CIRS, M1, T helper (Th)-1, Th-2, Th-17, T regulatory (Treg) profiles, and growth factors in depression and healthy controls. Culture supernatant of stimulated (5 μg/mL of PHA and 25 μg/mL of LPS) whole blood of 30 depressed patients and 20 controls was assayed for cytokines using the LUMINEX assay. The effects of three CBD concentrations (0.1 µg/mL, 1 µg/mL, and 10 µg/mL) were examined. Depression was characterized by significantly increased PHA + LPS-stimulated Th-1, Th-2, Th-17, Treg, IRS, CIRS, and neurotoxicity profiles. CBD 0.1 µg/mL did not have any immune effects. CBD 1.0 µg/mL decreased CIRS activities but increased growth factor production, while CBD 10.0 µg/mL suppressed Th-1, Th-17, IRS, CIRS, and a neurotoxicity profile and enhanced T cell growth and growth factor production. CBD 1.0 to 10.0 µg/mL dose-dependently decreased sIL-1RA, IL-8, IL-9, IL-10, IL-13, CCL11, G-CSF, IFN-γ, CCL2, CCL4, and CCL5, and increased IL-1β, IL-4, IL-15, IL-17, GM-CSF, TNF-α, FGF, and VEGF. In summary, in this experiment, there was no beneficial effect of CBD on the activated immune profile of depression and higher CBD concentrations can worsen inflammatory processes.
The potential role of cannabinoids in dermatology
The (poly) pharmacology of cannabidiol in neurological and neuropsychiatric disorders:molecular mechanisms and targets
DOI:10.3390/ijms22094876
URL
[本文引用: 1]
Cannabidiol (CBD), the major nonpsychoactive Cannabis constituent, has been proposed for the treatment of a wide panel of neurological and neuropsychiatric disorders, including anxiety, schizophrenia, epilepsy and drug addiction due to the ability of its versatile scaffold to interact with diverse molecular targets that are not restricted to the endocannabinoid system. Albeit the molecular mechanisms responsible for the therapeutic effects of CBD have yet to be fully elucidated, many efforts have been devoted in the last decades to shed light on its complex pharmacological profile. In particular, an ever-increasing number of molecular targets linked to those disorders have been identified for this phytocannabinoid, along with the modulatory effects of CBD on their cascade signaling. In this view, here we will try to provide a comprehensive and up-to-date overview of the molecular basis underlying the therapeutic effects of CBD involved in the treatment of neurological and neuropsychiatric disorders.
The anti-inflammatory effects of cannabidiol and cannabigerol alone,and in combination
外源水杨酸对镉胁迫下小报春幼苗生长及生理特性的影响
DOI:10.11733/j.issn.1007-0435.2020.05.020
[本文引用: 1]
为了探究叶面喷施外源水杨酸(Salicylic acid,SA)对小报春(Primula forbesii Franch.)镉(Cadmium,Cd)胁迫的缓解作用,本研究以小报春幼苗为试验材料,采用盆栽试验方法,在土壤Cd胁迫(150 mg·kg<sup>-1</sup>)下,测定不同浓度(0,10,100,1 000 μmol·L<sup>-1</sup>) SA处理下的小报春植株生长和生理特性指标。结果表明,低浓度SA (10,100 μmol·L<sup>-1</sup>)能够使小报春株高、冠幅和地上部生物量增加,促进光合作用;低浓度SA能降低过氧化氢和丙二醛含量,提高超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化物酶(Peroxidase,POD)和过氧化氢酶(Catalase,CAT)活性,促进可溶性糖和可溶性蛋白的产生;同时低浓度SA还能显著降低小报春幼苗叶片中的Cd含量,增加叶片中的钾和锌含量,降低钙和镁含量。而高浓度SA (1 000 μmol·L<sup>-1</sup>)缓解作用下降,甚至对植株生长产生抑制作用。本研究表明,100 μmol·L<sup>-1</sup>外源SA处理可以显著缓解150 mg·kg<sup>-1</sup>土壤Cd胁迫对小报春幼苗的毒害作用,并降低植株镉积累。
外源SA、GSH对Cd胁迫下绵毛水苏生理和生长的影响
DOI:10.11869/j.issn.100-8551.2021.01.0211
[本文引用: 1]
为探究不同外源物质对Cd胁迫下绵毛水苏生理和生长的缓解效果,采用盆栽试验研究叶面喷施不同浓度水杨酸(SA)(0.5、1.0、1.5、2.0 mmol·L<sup>-1</sup>)和谷胱甘肽(GSH)(0.1、0.2、0.3、0.4 mmol·L<sup>-1</sup>) 对300 mg·kg<sup>-1</sup> Cd胁迫下绵毛水苏幼苗生长、渗透调节物质、抗氧化酶活性以及Cd含量等的影响。结果表明,外源SA和GSH改善了植物的叶色,提高了叶面积和萌蘖能力,并增加了地下、地上部干物质积累量。除0.4 mmol·L<sup>-1</sup> GSH处理第3天外,外源SA和GSH处理均提高了可溶性糖、可溶性蛋白和游离脯氨酸等渗透调节物质的含量,最高增幅分别为16.06%、14.13%、311.39%(SA)和50.28%、12.77%、313.77%(GSH);此外,SA和GSH分别使抗氧化酶(SOD、POD和CAT)活性最高增幅依次达57.39%、38.51%、26.81%和85.00%、60.77%、50.21%。绵毛水苏吸收的Cd主要累积在根部,外源SA对Cd吸收无明显影响,而GSH抑制了Cd向地上部的转运。综上可知,SA和GSH对Cd胁迫下的绵毛水苏均具有缓解作用,以1.5 mmol·L<sup>-1</sup> SA和0.3~0.4 mmol·L<sup>-1</sup> GSH效果较佳。本研究结果对绵毛水苏应用于Cd污染土壤植物修复具有重要意义。
Salicylic acid-induced ROS production by mitochondrial electron transport chain depends on the activity of mitochondrial hexokinases in tomato (Solanum lycopersicum L.)
DOI:10.1007/s10265-019-01085-y
PMID:30758749
[本文引用: 1]
The growth regulator, salicylic acid (SA) plays an important role in the induction of cell death in plants. Production of reactive oxygen species (ROS) by mitochondrial electron transport chain (mtETC), cytochrome c (cyt c) release from mitochondria and loss of mitochondrial integrity can be observed during cell death execution in plant tissues. The aim of this work was to study the putative role of hexokinases (HXKs) in the initiation of cell death using tomato (Solanum lycopersicum L.) leaves and mitochondria isolated from plants exposed to a sublethal, 0.1 mM and a cell death-inducing, 1 mM concentrations of SA. Both treatments enhanced ROS and nitric oxide (NO) production in the leaves, which contributed to a concentration-dependent loss of membrane integrity. Images prepared by transmission electron microscopy showed swelling and disorganisation of mitochondrial cristae and vacuolization of mitochondria after SA exposure. Using post-embedding immunohistochemistry, cyt c release from mitochondria was also detected after 1 mM SA treatment. Both SA treatments decreased the activity and transcript levels of HXKs in the leaves and the total mtHXK activity in the mitochondrial fraction. The role of mitochondrial hexokinases (mtHXKs) in ROS and NO production of isolated mitochondria was investigated by the addition of HXK substrate, glucose (Glc) and a specific HXK inhibitor, N-acetylglucosamine (NAG) to the mitochondrial suspension. Both SA treatments enhanced ROS production by mtETC in the presence of succinate and ADP, which was slightly inhibited by Glc and increased significantly by NAG in control and in 0.1 mM SA-treated mitochondria. These changes were not significant at 1 mM SA, which caused disorganisation of mitochondrial membranes. Thus the inhibition of mtHXK activity can contribute to the mitochondrial ROS production, but it is not involved in NO generation in SA-treated leaf mitochondria suggesting that SA can promote cell death by suppressing mtHXK transcription and activity.
Physiological mechanisms for antagonistic interaction of manganese and aluminum in barley
DOI:10.1080/01904167.2019.1567767
[本文引用: 1]
In a previous study, we found that the combined addition of Al and Mn in the culture solution could alleviate the inhibition of barley growth by addition of Al or Mn alone. The current experiment was conducted in a greenhouse to investigate the physiological mechanisms of the antagonistic interaction using two barley genotypes, XZ16 (both Al and Mn tolerant) and ZU9 (both Al and Mn sensitive). The treatments consisted of three Al levels (0, 0.1, and 0.5 mM) and three Mn levels (0, 0.2, and 1.0 mM) and their combinations, and a completely randomized block design was used with three replications. The combined treatments had larger plant biomass, lower Al and Mn concentrations and accumulation in plant tissues, lower malondialdehyde content, and higher root ATPases activities, compared with Al or Mn alone treatment. The two genotypes had the similar trend in the antagonistic interaction, with ZU9 being more predominant than XZ16.
连续2年土壤砷胁迫对三七(Panax notoginseng)细胞膜透性和抗氧化酶活性的影响
Alleviation of lead-induced physiological,metabolic and ultramorphological changes in leaves of upland cotton through glutathione
DOI:10.1007/s11356-015-5959-4 URL [本文引用: 1]
AsA-GSH cycle and antioxidant enzymes play important roles in Cd tolerance of wheat
DOI:10.1007/s00128-018-2471-9
PMID:30353306
[本文引用: 1]
Wheat (Triticum aestivum L.) has relatively high tolerance to cadmium (Cd), but the underlying mechanisms are poorly understood. Growth and physiological parameters of wheat exposed to different Cd concentrations (0, 0.5, 5 and 50 µM) were characterized. The fresh weight, leaf chlorophyll and carotenoid concentrations and photosynthesis parameters did not differ among Cd treatments, suggesting relatively high Cd tolerance in wheat. However, the soluble sugar concentrations increased with the increasing Cd concentration and the soluble protein concentrations decreased in both shoots and roots, suggesting that the Cd application promoted nitrogen metabolism over carbon metabolism. In addition, the higher concentrations of MDA, GSH and AsA and activities of antioxidant enzymes (SOD, POD, and CAT) were observed in leaves and roots in the Cd50 treatment. Our results reveal that wheat can tolerate Cd by enhancing the antioxidant enzymes activities and increasing the concentration of ascorbate and glutathione.
Exogenous glutathione increased lead uptake and accumulation in Iris lactea var. chinensis exposed to excess lead
DOI:10.1080/15226514.2018.1460307 URL [本文引用: 1]
Cannabidiol protects against high glucose-induced oxidative stress and cytotoxicity in cardiac voltage-gated sodium channel
DOI:10.1111/bph.15020
PMID:32077098
[本文引用: 1]
Cardiovascular complications are the major cause of mortality in diabetic patients. However, the molecular mechanisms underlying diabetes-associated arrhythmias are unclear. We hypothesized that high glucose could adversely affect Na 1.5, the major cardiac sodium channel isoform of the heart, at least partially via oxidative stress. We further hypothesized that cannabidiol (CBD), one of the main constituents of Cannabis sativa, through its effects on Na 1.5, could protect against high glucose-elicited oxidative stress and cytotoxicity.To test these ideas, we used CHO cells transiently co-transfected with cDNA encoding human Na 1.5 α-subunit under control and high glucose conditions (50 or 100 mM for 24 hr). Several experimental and computational techniques were used, including voltage clamp of heterologous expression systems, cell viability assays, fluorescence assays and action potential modelling.High glucose evoked cell death associated with elevation in reactive oxygen species (ROS) right shifted the voltage dependence of conductance and steady-state fast inactivation, and increased persistent current leading to computational prolongation of action potential (hyperexcitability) which could result in long QT3 arrhythmia. CBD mitigated all the deleterious effects provoked by high glucose. Perfusion with lidocaine (a well-known sodium channel inhibitor with antioxidant effects) or co-incubation of Tempol (a well-known antioxidant) elicited protection, comparable to CBD, against the deleterious effects of high glucose.These findings suggest that, through its favourable antioxidant and sodium channel inhibitory effects, CBD may protect against high glucose-induced arrhythmia and cytotoxicity.© 2020 The British Pharmacological Society.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}