


土壤盐渍化是世界范围内严重的农业问题[1-2],是制约粮食生产的主要非生物胁迫之一[3]。据统计[4],我国约有25%的耕地受土壤盐度影响,且随着施肥量的增加,土壤次生盐渍化越来越严重,盐胁迫几乎影响到植物的整个生育期。盐度是限制农业生产力的主要非生物因素,通过影响植株形态、生理生化和代谢过程,包括种子萌发、水和养分吸收来阻碍其生长发育[5],造成作物光合减弱,使产量下降[6]。此外,盐胁迫影响植物的多种生理过程,包括光合作用、抗氧化酶活性、渗透剂[7]和激素机制[8]的调节。Bai等[9]研究发现,长期盐胁迫提高了燕麦根系超氧化物歧化酶(SOD)和过氧化物酶(POD)活性、丙二醛(MDA)和可溶性糖含量,降低了过氧化氢酶(CAT)活性。盐处理降低了燕麦的总叶面积、茎干重和叶片叶绿素含量,促进了燕麦叶片可溶性糖和脯氨酸的积累,且燕麦的耐盐性高于水稻(Oryza sativa L.)、小麦(Triticum aestium)和其他饲料作物[10]。因此,燕麦可作为改良盐渍土的先锋作物。
燕麦(Avena sativa L.)是全球第六大可用作食品、饲料的多用途谷物作物[11]。燕麦可在不同土壤和生长环境较差的地区种植,可用于改善盐碱土壤[12]。燕麦含有多种营养丰富和有益健康的物质,特别是酚类化合物[13],长期食用可有降血脂、降胆固醇和预防心血管疾病的功效[14]。前人[15-16]研究发现,盐胁迫不仅会影响燕麦苗期的生长、光合特性和脯氨酸含量,还会促进酚类物质的积累,提高植株的抗氧化能力,能够显著提高燕麦防御生物胁迫的能力。植物在不同生育时期对盐胁迫的响应机制明显不同[17],关于燕麦响应盐胁迫的研究大多数都集中在苗期,成熟期耐盐机制鲜见研究。因此,本研究选取2个耐盐性不同的燕麦品种作为试验材料,研究盐胁迫下燕麦成熟期生理指标、保护酶活性及酚类物质的变化规律,明确不同盐胁迫程度对成熟期燕麦生长发育和生理生化指标的影响。
1 材料与方法
1.1 试验材料
试验选取耐盐燕麦品种张燕7号(A)和盐敏感品种坝莜18号(B),均由张家口市农业科学院选育。
1.2 试验设计
试验用盐成分为NaCl和Na2SO4,按照质量比1:1混合。设4个盐胁迫浓度,分别为0.0%(对照)、0.2%、0.4%和0.6%,2个品种对应盐浓度处理分别用A0、A1、A2、A3、B0、B1、B2、B3表示,每个处理种植10盆,每盆播种30粒于装满配制基质(沙子:蛭石:陶粒=3:1:2)的塑料盆(高28 cm,直径22 cm),重复3次。培养温度25±2 ℃,光照/黑暗12 h/12 h。出苗3 d后计算发芽率,间苗留10株。在三叶期后,每隔7 d施加1/2 Hoagland营养液500 mL,在成熟期取叶片(从上到下数第1~2片叶),置于-80 ℃冰箱保存,用于测定相关生理指标,并收取种子。
1.3 测定项目与方法
1.3.1 生长指标
出苗率和株高:播种后3 d计算出苗率,在成熟期测定植株根部至穗顶距离(cm)。干、鲜重和叶片相对含水量测定:采收植株后用蒸馏水冲洗2~3次,擦干,测定单株鲜重(FW),后将植株在4 °C的去离子水中浸泡8 h后进行快速称重(TW),105 ℃烘箱干燥30 min,然后在80 ℃干燥至恒重后,测定单株干重(DW)。叶片相对含水量(RWC,%)=(FW-DW)/TW×100。
1.3.2 叶片叶绿素和光合指标
选取从顶部开始第3片,运用叶绿素测定仪(SPAD-502Plus)测定相对叶绿素含量(SPAD值),采用LI-6400便携式光合仪(LI-COR,美国)测定试验材料叶片的气孔导度(Gs)、净光合速率(Pn)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。
气孔限制值(Ls)=1-Ci/C0,式中,C0是大气CO2浓度(390 mmol/mol)。
1.3.3 酶活性和MDA含量
1.3.4 酚类物质和抗氧化酶活性
1.4 数据处理
采用SPSS 25.0软件对结果进行单因素方差分析(ANOVA)。
2 结果与分析
2.1 盐胁迫对燕麦生长的影响
由表1可知,2个燕麦品种株高随着盐浓度的增加呈逐渐降低的趋势,在0.2%~0.4%盐浓度下品种A较对照处理分别降低11.94%、26.05%,品种B分别降低13.10%、25.99%;0.6%盐浓度下,品种A和B较对照分别降低37.95%和33.29%,株高最低,可见盐胁迫抑制植株的生长,且不同品种株高在盐胁迫下响应不同。随着盐浓度的增加,2个品种的出苗率、单株鲜重和单株干重均呈下降趋势,且显著低于对照。0.6%盐浓度时,出苗率、单株鲜重和干重达到最低水平,品种A和B出苗率分别较对照降低21.88%和24.44%,鲜重分别降低53.33%和64.36%,干重分别降低60.76%和59.37%,说明盐胁迫显著降低燕麦的出苗率并抑制单株鲜重和干重,且品种B的出苗率明显低于品种A。随着盐浓度的升高,2个品种的RWC均呈先升高再降低的趋势,在盐浓度0.2%时达到最高,品种A和B分别较对照提高17.33%和12.20%,盐浓度在0.6%时,品种A和B叶片含水量达到最低值,较对照分别显著降低68.57%和71.95%。
表1 不同盐浓度对燕麦生长的影响
Table 1
| 处理 Treatment | 株高 Plant height (cm) | 出苗率 Emergence rate (%) | 单株干重 Dry weight per plant (g) | 单株鲜重 Fresh weight per plant (g) | 叶片相对含水量 RWC (%) |
|---|---|---|---|---|---|
| A0 | 84.53±1.70a | 96.05±1.03a | 1.58±0.05a | 7.72±0.20a | 75.06±6.01b |
| A1 | 74.43±1.22b | 91.12±1.58b | 1.25±0.05b | 6.56±0.34b | 88.32±2.23a |
| A2 | 62.51±1.40c | 82.33±1.02c | 0.93±0.07c | 5.82±0.03c | 68.44±2.58c |
| A3 | 52.45±1.05d | 75.35±1.56d | 0.62±0.01d | 3.00±0.08d | 27.07±2.77d |
| B0 | 79.11±2.65a | 90.11±0.67a | 1.28±0.05a | 6.65±0.09a | 82.88±2.30b |
| B1 | 68.75±2.42b | 84.49±1.59b | 1.09±0.05b | 5.37±0.08b | 92.09±2.04a |
| B2 | 58.55±2.11c | 80.71±1.06c | 0.73±0.02c | 4.69±0.14c | 76.65±2.87c |
| B3 | 52.78±1.09d | 68.66±1.55d | 0.52±0.01d | 2.37±0.08d | 23.37±3.08d |
不同小写字母表示差异显著(P < 0.05)。下同。
Different lowercase letters indicate significant difference (P < 0.05). The same below.
2.2 盐胁迫对燕麦叶片叶绿素含量和光合作用的影响
由图1可知,不同浓度盐胁迫对叶片SPAD和光合作用影响存在显著差异。2个品种的SPAD随着盐浓度的增加呈先升高后降低趋势。在0.2%和0.4%处理下,SPAD均高于对照,其中品种A的SPAD值较对照升高8.09%~12.55%,品种B升高3.30%~11.00%,品种A增幅均高于品种B。说明低浓度盐处理可促进叶绿素形成,过高的盐浓度破坏了叶片中叶绿素结构,抑制了叶绿素形成。在0.2%盐浓度下,2个品种的Pn和Ci都显著高于对照,Pn分别升高10.46%和12.01%,Ci分别提高17.81%和16.06%;在0.4%~0.6%浓度盐胁迫下,2个品种的Pn和Ci值显著低于对照水平,品种A的Pn和Ci减幅明显低于品种B。随着盐浓度的升高,Tr为先降低再升高的趋势,在0.4%盐处理下,品种A和B的Tr达到最高值,显著高于对照,分别提高13.63%和10.65%。随着盐浓度的升高,Gs均呈降低的趋势,在盐浓度0.4%~0.6%下,Gs显著低于对照水平,品种A降幅为9.00%和37.83%,B降幅为20.51%和47.43%,品种B减幅较大。除对照外,2个品种的Ls随着盐浓度的增加均逐渐提高,在0.6%浓度盐处理下Ls达到最高,品种A较B高,说明气孔因素是盐处理下光合速率降低的主要原因。
图1
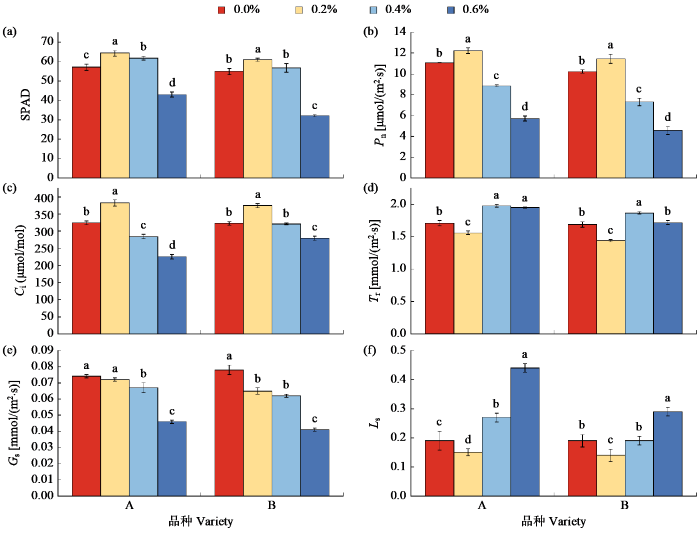
图1
不同盐浓度对SPAD和光合指标的影响
不同小写字母表示处理间差异显著(P < 0.05),下同。
Fig.1
Effects of different salt concentrations on SPAD and photosynthetic indices
Different lowercase letters indicate significant differences between treatments (P < 0.05), the same below.
2.3 盐胁迫对燕麦抗氧化酶活性的影响
SOD活性直接影响了作物活性氧的清除能力,与作物抗盐性息息相关。表2显示,随着盐浓度的升高,品种A和B的SOD和POD活性都呈现先升高再降低的趋势,在0.2%盐浓度下,品种A和B的SOD活性显著高于对照,分别增加了51.06%和7.06%,POD分别增加29.55%和52.38%,在0.6%浓度盐处理下,品种A和B的SOD活性显著高于对照,分别增加了19.60%和20.50%。品种A和B分别在0.2%和0.4%盐浓度下SOD活性达到最高峰,且品种A的SOD活性增幅较高。随着盐胁迫浓度的升高,品种A和B的CAT活性和MDA含量呈升高趋势,都显著高于对照,在0.6%盐浓度CAT活性分别较对照升高109.72%和36.97%,MDA含量分别升高18.41%和62.60%。说明盐胁迫下耐盐品种具有较强的CAT和SOD活性,燕麦通过升高CAT和SOD活性来提高耐盐性。
表2 不同盐浓度对燕麦抗氧化酶活性和MDA含量的影响
Table 2
| 处理 Treatment | SOD活性 SOD activity [U/(min·g)] | POD活性 POD activity [U/(min·g)] | CAT活性 CAT activity [U/(min·g)] | MDA含量 MDA content (nmol/g) |
|---|---|---|---|---|
| A0 | 7928.33±44.88d | 2.03±0.02b | 2.88±0.02d | 31.11±0.67c |
| A1 | 11 976.67±29.87a | 2.63±0.14a | 4.57±0.08c | 31.41±0.60c |
| A2 | 9890.33±63.26b | 1.64±0.14c | 5.33±0.08b | 33.95±0.06b |
| A3 | 9482.33±89.51c | 0.99±0.02d | 6.04±0.05a | 36.84±0.05a |
| B0 | 7255.33±42.10d | 1.68±0.18c | 4.03±0.03d | 53.55±0.69d |
| B1 | 7767.67±63.01c | 2.56±0.08a | 4.61±0.04c | 61.26±1.01c |
| B2 | 8890.67±20.11a | 2.05±0.05b | 5.05±0.02b | 78.51±0.86b |
| B3 | 8742.33±21.96b | 1.20±0.08d | 5.52±0.06a | 87.07±1.48a |
2.4 盐胁迫对燕麦籽粒酚类物质及抗氧化能力的影响
由图2可知,盐胁迫显著提高了燕麦籽粒中的总酚和总黄酮含量,在盐浓度0.6%时,燕麦籽粒总酚含量达到最高水平,且较对照分别提高32.54%和21.37%。在盐浓度0.4%时,总黄酮含量达到最大值,品种A和B较对照分别提高36.72%和21.87%。说明随着盐浓度的提高,燕麦籽粒中的总酚和总黄酮含量会显著增加,而且品种A明显高于品种B。盐胁迫增加了2个品种的ABTS和DPPH抗氧化能力,在盐浓度0.6%时,2个品种籽粒的ABTS和DPPH抗氧化能力达到最高,品种A和B的ABTS值最高分别为81.73%和86.31%,且较对照分别提高23.29%和32.52%;DPPH最高分别为80.68%和77.79%,且较对照分别提高29.05%和39.73%。品种A的ABTS抗氧化能力略低于品种B,而DPPH高于品种B。
图2
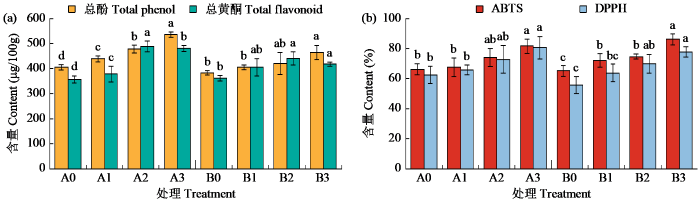
图2
不同盐浓度对燕麦总酚、总黄酮及抗氧化能力的影响
Fig.2
Effects of different salt concentrations on total phenols, total flavonoid and antioxidant capacity of oat
2个品种的芦丁、木樨草素和芹菜素含量整体随着盐胁迫浓度的增加逐渐提高(表3),在盐浓度0.6%时,芦丁、木樨草素和芹菜素含量显著高于其他处理(除A2处理),品种A较对照分别提高330.05%、118.79%、69.06%,品种B分别提高291.69%、298.86%、165.43%,品种B的含量高于品种A。盐浓度在0.4%时,品种A籽粒中咖啡酸、对香豆酸、阿魏酸、2,3-二羟基苯甲酸、对羟基苯甲酸和绿原酸含量较对照提高30.32%~271.68%。品种B除了2,3-二羟基苯甲酸在盐浓度0.4%达到最高水平外,其他5种酚类化合物在盐浓度0.6%时含量达到最高水平,分别较对照提高56.94%~ 534.61%。品种A和B籽粒中香草酸的含量分别在盐浓度0.6%和0.4%时达到最高水平,品种A最高含量为164.85 μg/100 g,较对照显著提高272.71%;但在0.6%盐浓度时,品种B香草酸含量达到最低,说明随着盐浓度的提高,品种B籽粒中的香草酸含量积累受到抑制。
表3 不同盐浓度对燕麦单体酚的影响
Table 3
| 处理 Treatment | 木犀草素 Luteolin | 芹菜素 Apigenin | 2,3-二羟基苯甲酸 2,3-dihydroxybenzoic acid | 对羟基苯甲酸 P-hydroxybenzoic acid | 绿原酸 Chlorogenic acid |
|---|---|---|---|---|---|
| A0 | 58.48±1.98d | 147.13±6.31d | 83.70±0.80d | 260.08±1.68d | 160.33±0.29b |
| A1 | 74.68±0.66c | 153.66±1.78c | 125.82±7.11c | 320.55±19.95c | 136.22±0.19d |
| A2 | 136.83±1.26a | 205.56±1.91b | 155.82±5.69b | 481.44±5.71b | 143.91±1.28c |
| A3 | 127.95±1.69b | 248.74±2.23a | 197.49±6.30a | 521.74±2.75a | 205.78±0.33a |
| B0 | 44.09±0.76d | 115.78±0.50d | 71.35±2.45d | 391.31±2.80b | 86.32±1.81d |
| B1 | 56.66±2.23c | 193.40±2.51c | 97.35±6.35c | 412.75±7.59b | 105.22±1.21c |
| B2 | 99.72±4.65b | 239.52±4.57b | 183.89±6.12a | 812.49±29.61a | 126.09±2.92b |
| B3 | 175.86±1.59a | 307.31±8.96a | 168.02±1.07b | 823.11±24.61a | 154.27±1.84a |
3 讨论
3.1 盐胁迫对燕麦生长和光合指标的影响
盐胁迫下不同基因型燕麦出苗率有较大差异,种子发芽率、发芽势以及活力指数随着盐浓度的增加均呈下降趋势[23]。本研究中,株高和种子萌发率随着盐浓度的增加略有下降,与Zhang等[24]研究结果一致。说明在种子萌发过程中,由于膜的选择性效率降低,可能会导致达到毒害浓度的离子进入细胞原生质,进而影响种子发芽、出苗。但Foolad等[25]研究发现,在盐胁迫条件下,种子萌发速度是不变的,可能存在耐胁迫基因或特殊的生理机制。本文中鲜重、干重和株高均呈降低趋势且株高在0.6%盐浓度下达到最低水平,与徐正辉[26]研究结果一致。在本试验中,在盐胁迫下品种A和B的株高降低程度差异不大,有可能由于在整个生长过程中试验条件和盐胁迫浓度等差异,在盐胁迫下株高有所降低,但是差异不大。RWC均为先升高再降低的趋势,在高盐浓度下,RWC达到最低值,发现过量的盐会导致种子对水分的吸收减少,验证了高浓度盐胁迫严重抑制植株的生长发育。
盐度可以通过抑制叶绿素合成或加速叶绿素降解进而影响叶绿素含量,从而影响植物的光合作用和干物质的积累。随盐浓度增加,耐盐品种A叶绿素含量高于盐敏感品种B,这与王旭明等[27]在水稻和Sapre等[28]在燕麦上研究结果一致,可能是叶绿体结构在盐胁迫下受到了伤害。盐胁迫会抑制光合作用,降低光合速率,最终影响产量。本研究表明,随着盐浓度的增加,燕麦的Pn、Ci、Gs显著降低,与王苗苗等[29]研究结果一致。光合作用的降低很可能是因为Na+影响气孔关闭及光合相关基因表达,使光合速率减缓,相关指标总体呈下降趋势,从而减轻了盐胁迫对燕麦光合器官的损伤,盐敏感品种较耐盐品种下降更为明显,与孙璐等[30]在高粱上研究结果一致,本文中光合参数与叶绿素含量变化趋势基本保持一致,说明随着盐胁迫浓度的升高,叶绿素含量降低会进一步影响燕麦光能的吸收和转化,进而导致光合利用率降低,是最终导致燕麦产量降低的主要原因。
3.2 盐胁迫对燕麦酶活性、酚类物质及抗氧化能力的影响
盐胁迫下植物保护酶活性会发生变化,如活性氧的产生和清除平衡被破坏,保护酶活性被抑制。SOD是植物体内氧代谢的关键酶,能催化歧化反应,其活性的高低影响植物体内O2和H2O2的含量。MDA含量反映膜过氧化的强度,其含量高低和细胞膜受损情况成正比,与抗逆能力成反比[31]。Bai等[9]研究发现,盐胁迫后燕麦根系中的SOD和POD活性比对照显著增加。本研究发现,随着盐浓度的升高,2个品种的SOD和POD活性都呈现先升高再降低的趋势,与Wu等[32]研究结果一致,说明低盐浓度可引起燕麦叶片SOD和POD活性的升高,能清除自由基,防止膜的过氧化和细胞受到伤害,是盐胁迫的防御反应。而在高浓度盐胁迫下,以SOD为主的细胞保护系统活性降低,导致原生质膜脂质和膜结构发生变化,相对电导率和MDA含量升高[33]。本研究发现随着盐胁迫浓度升高,2个品种的CAT活性和MDA含量呈升高趋势,说明高盐度加剧了膜脂过氧化程度并造成细胞膜系统损伤,这与刘文瑜等[34]在藜麦中的研究结果一致。低盐胁迫下可诱导保护酶活性升高,并有效清除活性氧,进而降低MDA含量。通过SOD、POD和CAT等抗氧保护酶系统的协同作用,不断产生和排出活性氧,调节植物正常代谢水平不受破坏[35]。刘凤歧等[36]研究了不同燕麦对盐胁迫的生理响应,发现耐盐品种的抗氧化酶活性和渗透调节物质高于盐敏感品种,表明耐盐品种倾向于保持较高水平的活性氧清除活性[37]。
4 结论
盐胁迫抑制了燕麦生长、光合指标和酶活性,但使燕麦积累较高的酶活性、酚类物质含量及抗氧化能力,从而抵御一定程度的盐胁迫,保证其正常生长,盐胁迫对盐敏感品种坝莜18号抑制作用大于耐盐品种张燕7号。低盐浓度(0.2%~0.4%)胁迫下,Pn、Ci、SOD活性、POD活性、SPAD和Tr有所提高,但在高盐浓度(0.6%)下,燕麦籽粒的酚类物质含量及抗氧化能力显著提高。
参考文献
Salinity differentially affects growth and ecophysiology of two mastic tree (Pistacia lentiscus L.) accessions
Protection of halophytes and their uses for cultivation of saline-alkali soil in China
Integrated physiologic,proteomic,and metabolomic analyses of Malus halliana adaptation to saline-alkali stress
Rapid and tissue-specific changes in ABA and in growth rate in response to salinity in barley leaves
The addition of 100 mM NaCl to the root medium of barley plants caused the rapid cessation of elongation of the growing leaf three, followed by a sudden resumption of growth during the following hour. The idea that resumption of growth is preceded and mediated by rapid and tissue-specific changes in ABA concentration and by changes in transpiration was tested. Leaf elongation velocity was recorded continuously using linear variable displacement transducers (LVDT), ABA was determined by immunoassay, and transpiration and stomatal conductivity were measured gravimetrically and by porometry, respectively. Within 10 min following addition of salt, ABA increased 6-fold in the distal portion of the leaf elongation zone; in the proximal portion, ABA accumulated with a delay. In the portion of the growing blade that had emerged ABA increased 3-fold and remained elevated during the following 20 min. This preceded a decrease in transpiration and stomatal conductivity, which, in turn, coincided with growth resumption. Twenty hours following the addition of salt, the ABA concentrations had returned to the level before stress. Leaf elongation velocity was still reduced. It is concluded that NaCl causes a rapid increase in ABA in the transpiring portion of the growing leaf. This leads to a decrease in transpiration. As a result, xylem water potential is expected to rise. The moment that the water potential gradient between the xylem and the peripheral cells in the growth zone favours water uptake again into the latter, leaf elongation resumes. The results suggest that ABA causes different responses in different leaf regions, all aimed at promoting the resumption of leaf growth.
Proteomic analysis of salt-responsive proteins in oat roots (Avena sativa L.)
Haplotype-based genotyping-by-sequencing in oat genome research
DOI:10.1111/pbi.12888
PMID:29345800
[本文引用: 1]

In a de novo genotyping-by-sequencing (GBS) analysis of short, 64-base tag-level haplotypes in 4657 accessions of cultivated oat, we discovered 164741 tag-level (TL) genetic variants containing 241224 SNPs. From this, the marker density of an oat consensus map was increased by the addition of more than 70000 loci. The mapped TL genotypes of a 635-line diversity panel were used to infer chromosome-level (CL) haplotype maps. These maps revealed differences in the number and size of haplotype blocks, as well as differences in haplotype diversity between chromosomes and subsets of the diversity panel. We then explored potential benefits of SNP vs. TL vs. CL GBS variants for mapping, high-resolution genome analysis and genomic selection in oats. A combined genome-wide association study (GWAS) of heading date from multiple locations using both TL haplotypes and individual SNP markers identified 184 significant associations. A comparative GWAS using TL haplotypes, CL haplotype blocks and their combinations demonstrated the superiority of using TL haplotype markers. Using a principal component-based genome-wide scan, genomic regions containing signatures of selection were identified. These regions may contain genes that are responsible for the local adaptation of oats to Northern American conditions. Genomic selection for heading date using TL haplotypes or SNP markers gave comparable and promising prediction accuracies of up to r = 0.74. Genomic selection carried out in an independent calibration and test population for heading date gave promising prediction accuracies that ranged between r = 0.42 and 0.67. In conclusion, TL haplotype GBS-derived markers facilitate genome analysis and genomic selection in oat.© 2018 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Comparative biomass production of six oat varieties in a saline soil ecology
Subgenome-specific assembly of vitamin E biosynthesis genes and expression patterns during seed development provide insight into the evolution of oat genome
DOI:10.1111/pbi.12571
PMID:27135276
[本文引用: 1]
Vitamin E is essential for humans and thus must be a component of a healthy diet. Among the cereal grains, hexaploid oats (Avena sativa L.) have high vitamin E content. To date, no gene sequences in the vitamin E biosynthesis pathway have been reported for oats. Using deep sequencing and orthology-guided assembly, coding sequences of genes for each step in vitamin E synthesis in oats were reconstructed, including resolution of the sequences of homeologs. Three homeologs, presumably representing each of the three oat subgenomes, were identified for the main steps of the pathway. Partial sequences, likely representing pseudogenes, were recovered in some instances as well. Pairwise comparisons among homeologs revealed that two of the three putative subgenome-specific homeologs are almost identical for each gene. Synonymous substitution rates indicate the time of divergence of the two more similar subgenomes from the distinct one at 7.9-8.7 MYA, and a divergence between the similar subgenomes from a common ancestor 1.1 MYA. A new proposed evolutionary model for hexaploid oat formation is discussed. Homeolog-specific gene expression was quantified during oat seed development and compared with vitamin E accumulation. Homeolog expression largely appears to be similar for most of genes; however, for some genes, homoeolog-specific transcriptional bias was observed. The expression of HPPD, as well as certain homoeologs of VTE2 and VTE4, is highly correlated with seed vitamin E accumulation. Our findings expand our understanding of oat genome evolution and will assist efforts to modify vitamin E content and composition in oats.Published 2016. This article is a U.S. Government work and is in the public domain in the USA. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Polyamine catabolism influences antioxidative defense mechanism in shoots and roots of five wheat genotypes under high temperature stress
Total phenolics,flavonoids and antioxidant activity of tropical fruit pineapple
Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts
Effects of mixed salt stress on germination percentage and protection system of oat seedling
Relationships between cold- and salt-tolerance during seed germination in tomato: germplasm evaluation
NaCl胁迫对不同耐盐性水稻某些生理特性和光合特性的影响
DOI:10.3969/j.issn.1000-2561.2019.05.008
[本文引用: 1]
选取较耐盐的水稻品种‘HH11’、‘JX99’和盐敏感水稻品种‘YSXD’,设置6个土壤NaCl浓度处理(0、1、2、3、4 g/kg),在防雨棚下盆栽并培育至孕穗期,分析NaCl胁迫对不同耐盐性水稻生理生化及光合特性的影响,结果表明:(1)NaCl胁迫抑制水稻的生长,表现为随着NaCl浓度增加,水稻的株高逐渐降低,但在3~4 g/kg土壤含盐量下耐盐水稻的株高显著高于盐敏感水稻品种。(2)耐盐水稻和盐敏感水稻的可溶性总糖对NaCl胁迫的响应差异明显,在1~4 g/kg NaCl胁迫下,盐敏感水稻叶片可溶性总糖显著降低,但是耐盐水稻可溶性总糖大量合成并积累,并且显著高于盐敏感品种。(3)NaCl浓度增加迫使水稻叶片丙二醛不断积累,导致细胞膜透性逐渐增大,但是耐盐水稻丙二醛的积累量较少,细胞膜受盐害程度显著小于盐敏感水稻。(4)NaCl胁迫抑制了水稻光合速率,但不同的NaCl浓度下导致水稻品种光合速率下降的原因各有差异,其中在0~1 g/kg NaCl胁迫下盐敏感水稻光合速率降低是非气孔因素导致的,而在2~4 g/kg NaCl胁迫下是由气孔因素造成的;0~2 g/kg盐浓度下耐盐水稻HH11的光合速率降低是非气孔因素导致的,3~4 g/kg盐浓度处理是由气孔因素导致的;气孔因素是0~4 g/kg盐处理耐盐水稻JX99的光合速率降低的主要原因。(5)盐胁迫下耐盐水稻的叶片蒸腾速率显著降低,并且显著低于盐敏感水稻,相反水分利用效率和气孔限制值却明显升高,并且显著高于盐敏感水稻品种,表明盐浓度增加迫使耐盐水稻气孔阻力增大,减少水分的流失,抑制了蒸腾速率,使耐盐水稻叶片保持较高的水势;同时提高了叶片水分利用效率,碳同化效率提高,以满足耐盐水稻正常代谢生理需求,进行正常生命活动。
Klebsiella sp. confers enhanced tolerance to salinity and plant growth promotion in oat seedlings (Avena sativa)
盐胁迫对高粱幼苗光合作用和荧光特性的影响
DOI:10.3864/j.issn.0578-1752.2012.16.005
[本文引用: 1]
【目的】研究盐胁迫对不同高粱品种幼苗光合作用及叶绿素荧光参数的影响,为高粱栽培管理、耐盐品种的选育及耐盐胁迫人工调控提供理论依据。【方法】以高粱耐盐品种(辽杂15号)和盐敏感品种(龙杂11号)为材料,人工气候箱内营养液培养,湿度60%,光照/黑暗为12 h/12 h,光照强度为134 μmol•m-2•s-1,昼/夜温度为28℃/25℃。从3叶期开始进行NaCl处理(NaCl浓度为0、50、100、150、200 mmol•L-1),研究盐胁迫下高粱幼苗光合作用和叶绿素荧光参数的变化。【结果】低浓度NaCl(50 mmol•L-1)胁迫可以增加叶绿素含量,但是高浓度NaCl(100—200 mmol•L-1)胁迫明显降低叶绿素含量;盐胁迫使净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、最大荧光(Fm)、Fv/Fo、Fv/Fm、Fv′/Fm′、光化学猝灭系数(qP)下降,初始荧光(Fo)和非光化学猝灭系数(NPQ)提高,低浓度盐胁迫(50 mmol•L-1 NaCl)使胞间CO2浓度(Ci)降低,高浓度则相反。辽杂15号受盐胁迫的影响程度小于龙杂11号,表现出较好的耐盐性。【结论】50 mmol•L-1浓度的低盐胁迫对高粱幼苗的影响不明显,光合速率下降的主要原因是气孔限制;100—200 mmol•L-1浓度的高盐胁迫对高粱幼苗有很大影响,引起光合速率下降的主要原因是非气孔限制。盐胁迫下,耐盐品种能有效保护内部的光合机构,增强对盐胁迫的适应性。
Comparative transcriptional profiling and physiological responses of two contrasting oat genotypes under salt stress
DOI:10.1038/s41598-018-34505-5
PMID:30389990
[本文引用: 1]
Salinity is one of the major abiotic factors that affect productivity in oat. Here, we report a comparison of the transcriptomes of two hexaploid oat cultivars, 'Hanyou-5' and 'Huazao-2', which differ with respect to salt tolerance, in seedlings exposed to salt stress. Analysis of the assembled unigenes from the osmotically stressed and control libraries of 'Hanyou-5' and 'Huazao-2' showed that the expression of 21.92% (36,462/166,326) of the assembled unigenes was differentially regulated in the two cultivars after different durations of salt stress. Bioinformatics analysis showed that the main functional categories enriched in these DEGs were "metabolic process", "response to stresses", "plant hormone signal transduction", "MAPK signalling", "oxidative phosphorylation", and the plant-pathogen interaction pathway. Some regulatory genes, such as those encoding MYB, WRKY, bHLH, and zinc finger proteins, were also found to be differentially expressed under salt stress. Physiological measurements also detected significant differences in the activities of POD (76.24 +/- 1.07 Vs 81.53 +/- 1.47 U/g FW) in the two genotypes in response to osmotic stress. Furthermore, differential expression of 18 of these genes was successfully validated using RNA-seq and qRT-PCR analyses. A number of stress-responsive genes were identified in both cultivars, and candidate genes with potential roles in the adaptation to salinity were proposed.
4种燕麦对NaCl胁迫的生理响应及耐盐性评价
DOI:10.11686/cyxb20150122
[本文引用: 1]
为研究4份燕麦坝莜一号、白燕2号、白燕6号和白燕7号的耐盐程度及其在盐胁迫处理下的生理反应规律,在水培条件下,分别用0,34.2,68.4,102.6,136.8,171.0 mmol/L浓度NaCl溶液模拟盐胁迫处理4种燕麦种子,观察记录各处理的盐害情况,分别测定了胁迫后的种子发芽势、发芽率、株高和根长,以及室温培养的燕麦幼苗叶片过氧化物酶(POD)、超氧化物歧化酶(SOD)和可溶性蛋白含量,分析了不同胁迫程度下各生理指标的变化情况,并且对4种燕麦的耐盐性进行了综合评价。结果表明,NaCl胁迫对4种燕麦种子的发芽势和发芽率均产生显著影响,对株高和根长产生了抑制作用,且均随盐浓度的增大呈逐渐降低的趋势;燕麦幼苗叶片POD活性、SOD活性、可溶性蛋白含量均在102.6 mmol/L 盐浓度下达到最大值,而且都随盐浓度的增大呈先升高后降低的趋势,表明燕麦具有较高的耐盐能力,在盐胁迫下可采取自我保护机制以适应盐胁迫,4种燕麦的耐盐阈值为102.6 mmol/L盐浓度。综合评价,白燕6号耐盐性较好,白燕2号具有高盐耐受性的潜力,这两个燕麦品种可以为耐盐基因挖掘及燕麦耐盐品种的选育提供材料。
Phenolic composition and antioxidant activity of colored oats
DOI:S0308-8146(18)31006-9
PMID:30064743
[本文引用: 2]
Phenolic composition and antioxidant capacity of 20 oat genotypes differing in hull color were investigated. Phenolic aldehydes, phenolic acids, avenanthramides and mono-, and diglycerides were identified in the soluble phenolic fraction of the genotypes. The bound phenolic fraction proved to be less diverse with phenolic aldehydes, phenolic acids and a ferulic acid dehydrodimer detected. Investigating the scavenging capacity of the hull and groat toward 2,2-diphenyl-1-picrylhydrazyl (DPPH), an increased antioxidant activity (AOA) of hull compared to groats and a color dependence of the hull AOA could be observed. Principal component analysis on the determined variables revealed that the black-hulled samples were different from the white ones due to their increased phenolic content detected in the hull. However, reddish-hulled varieties were grouping with the accessions of the other colored groups. In addition, a distinction between spring and winter cultivars was also observed.Copyright © 2018 Elsevier Ltd. All rights reserved.

{kind=link}
{kind=link}
{kind=link}
{kind=link}