


在植物吸收氮素过程中,硝态氮必须转化成铵态氮后才能被植物同化,再通过氨基化、转氨基、酰胺化作用后合成蛋白质。在无机氮转化为有机氮的过程中,植物体内的谷氨酰胺合成酶(glutamine synthetase,GS)、谷氨酸脱氢酶(glutamate dehydrogenase,GDH)和硝酸还原酶(nitrate reductase,NR)对氮代谢关键酶起重要作用[5],直接影响植物对氮素的吸收和利用。硝酸还原酶是植物体内无机氮转化为有机氮的限速酶,其活性高低直接影响氮代谢水平[6]。研究[7-8]认为,适宜的氮肥来源和施用量可以显著提高氮代谢相关酶的活性。甜菜是高耗氮作物,在生产过程中往往需要多施氮肥来提高甜菜块根产量及其他生物量指标。然而,过量施氮使甜菜体内氮代谢旺盛,同时,过量的硝酸根离子消耗糖代谢中的碳水化合物形成氨基酸、酰胺等,使有害氮含量增加[9]。
本研究探讨了6个硝态氮、铵态氮配比水平下甜菜的表型、可溶性蛋白含量以及叶绿素含量等的差异,分析了在不同氮素形态配比的培养条件下,甜菜体内NR、GS和GDH 3种氮同化关键酶的活性和相关的调控机制,旨在探讨不同氮素形态配比对甜菜生长状况的影响以及氮同化的相关调控机制与甜菜生长之间的关系,为揭示不同氮素形态对甜菜生长以及相关的代谢过程提供科学依据。
1 材料与方法
1.1 试验材料
以甜菜品种KWS8138(德国KWS公司,二倍体单粒种)为试验材料。
1.2 试验设计
将大小均匀、无明显病虫害的种子在2%福美双浸泡一夜之后,用清水清洗,用盛有蛭石的培养槽进行10 d的育苗培养。当2片子叶完全展开后,移栽到铵硝比分别为0:5、4:1、3:2、2:3、1:4、5:0改良的Hoagland营养液中,于2022年9-12月在黑龙江大学国家甜菜种质中期库智能人工气候室内进行,昼、夜温度分别为25和18 ℃,光照强度200 µmol/(m2·s),光照周期为黑暗处理14 h,光照处理10 h。分别于甜菜长出2片真叶后的14(幼苗期)、36(叶丛快速增长期)、60 d(块根、糖分增长期)对甜菜各生理指标进行测定。
1.3 测定项目与方法
样本表型主要包括茎粗、株高、根长、根鲜重、叶鲜重、根干重、叶干重、根饱和重。随机取样,每一个处理取3株。使用游标卡尺测量茎粗;使用直尺测量株高、根长;将每株甜菜幼苗的叶部放入玻璃罐中,加水放置4 h,擦干水分称量根鲜重、叶鲜重、叶饱和重;在80 ℃烘干样本3 h后用电子天平分别称量根干重、叶干重。
采用95%乙醇提取法测定叶绿素含量,具体为称取0.5 g叶片置于10 mL离心管中,加入10 mL乙醇,浸泡48 h,取上清液,使用多功能酶标仪分别测量在665(A665)和649 nm(A649)的最大吸收峰处的吸光值,通过公式计算可得。
式中,A665、A649分别为叶绿素提取液在波长665、649 nm下的吸光度,Ca表示叶绿素a的含量,Cb表示叶绿素b含量,Ca+b表示叶绿素总含量。
使用BCA蛋白定量试剂盒(江苏康为世纪生物科技股份有限公司,产品编号为BL521A)测定可溶性蛋白含量(mg/L)。使用硝酸还原酶(NR)活性测定试剂盒(南京建成生物工程研究所,产品编号为A096-1-2)测定NR活性。使用GS测定试剂盒(南京建成生物工程研究所,产品编号为A047-1-1)测定GS活性。使用GDH测试盒(南京建成生物工程研究所,产品编号为A1125-1-1)测定GDH活性。
1.4 数据处理
使用Excel 2019处理数据,使用SPSS 26.0进行单因素方差性分析、相关性分析和主成分分析,使用Origin 2021作图。
2 结果与分析
2.1 不同氮素形态配比下甜菜的表型差异
由图1可知,不同氮素形态配比对甜菜的茎粗、株高、根长、根鲜重、叶鲜重、根干重、叶干重、叶饱和重有显著影响。多数植物对不同氮素的形态喜好不尽相同,甜菜是一种喜硝作物,对硝态氮的吸收要比铵态氮容易的多。在纯硝态氮供应下,甜菜的茎粗、根干重、根鲜重在整个生长时期中表现最好,在14 d表现的尤为明显。根据茎粗、根鲜重、叶鲜重、根干重、叶干重、叶饱和重等指标发现,铵态氮的增加对甜菜生长没有明显的促进作用,在纯铵态氮的供应下,这些指标均低于其他处理。根长和株高随着铵态氮的增加没有明显降低。
图1
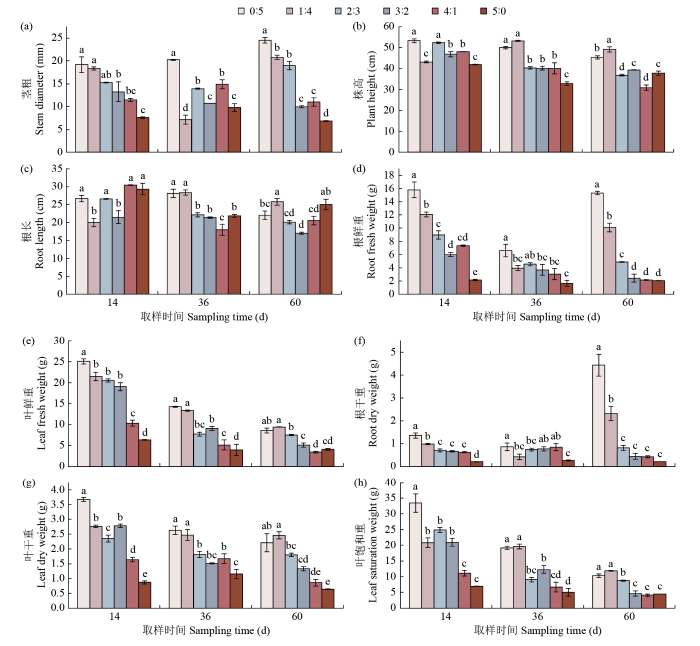
图1
不同氮素形态配比下甜菜生长的各项指标
不同小写字母表示同一时期内不同处理间在P < 0.05有显著差异,下同。
Fig.1
Each growth index of sugar beet under different nitrogen form ratios
Different lowercase letters indicate significant differences at P < 0.05 between treatments in the same period, the same below.
2.2 不同氮素形态配比对甜菜各生理生化指标的影响
2.2.1 对叶绿素含量的影响
由表1可知,与纯铵态氮供应相比,不同的铵硝配比显著提高了甜菜体内的叶绿素含量。处理14 d后,叶绿素含量随着铵态氮的施加呈现降低―升高―再降低的趋势,并且在铵硝比为3:2时含量达到最高。在36和60 d时,叶绿素含量在铵硝比为2:3时达到最高。根据这些表现来看,合理的铵硝配比能够显著提高甜菜的叶绿素含量。
表1 不同氮素形态配比对叶绿素含量的影响
Table 1
| 处理 Treatment | 取样时间Sampling time | ||
|---|---|---|---|
| 14 d | 36 d | 60 d | |
| 0:5 | 29.995±0.341b | 7.146±0.029cd | 6.699±0.231c |
| 1:4 | 29.868±0.090ab | 4.381±0.369d | 4.237±0.361d |
| 2:3 | 22.935±0.028c | 10.956±0.005a | 10.490±0.001a |
| 3:2 | 31.388±0.354a | 7.088±0.022c | 6.692±0.161c |
| 4:1 | 24.226±0.045c | 8.795±0.184b | 8.477±0.185b |
| 5:0 | 6.349±0.025d | 3.484±0.342e | 3.415±0.349e |
不同小写字母表示同一时期内不同处理在P < 0.05有显著差异,下同。
Different lowercase letters indicate significant differences at P < 0.05 between treatments in the same period, the same below.
2.2.2 对可溶性蛋白含量的影响
根据表2可知,随着处理时间的增长,可溶性蛋白含量呈先升高后降低的趋势。甜菜地下部可溶性蛋白含量明显低于叶片。与纯铵态氮供应和纯硝态氮供应相比,不同铵硝比处理下14和36 d的可溶性蛋白含量均有增加。随着铵态氮比例的增加,可溶性蛋白含量呈现先升高后降低的趋势。处理36 d后,当铵硝比为4:1时叶片的可溶性蛋白含量最高,为3.430 mg/L,证明了铵态氮和硝态氮同时施加可以提高植物体内的可溶性蛋白含量。
表2 不同氮素形态配比对甜菜可溶性蛋白含量的影响
Table 2
| 部位 Part | 处理 Treatment | 取样时间Sampling time | ||
|---|---|---|---|---|
| 14 d | 36 d | 60 d | ||
| 叶Leaf | 0:5 | 0.646±0.022c | 2.470±0.020bc | 1.885±0.135ab |
| 1:4 | 0.984±0.063a | 3.046±0.245ab | 1.270±0.073cd | |
| 2:3 | 0.867±0.015b | 3.136±0.098a | 1.733±0.180b | |
| 3:2 | 0.952±0.008a | 2.901±0.570abc | 2.265±0.341a | |
| 4:1 | 0.867±0.015b | 3.430±0.124a | 1.568±0.201bc | |
| 5:0 | 0.697±0.027c | 2.414±0.107c | 1.059±0.179d | |
| 根Root | 0:5 | 0.230±0.010cd | 0.429±0.008a | 0.244±0.001a |
| 1:4 | 0.243±0.013bc | 0.303±0.041b | 0.237±0.007a | |
| 2:3 | 0.250±0.005abc | 0.213±0.002c | 0.255±0.018a | |
| 3:2 | 0.271±0.006a | 0.242±0.034c | 0.226±0.001a | |
| 4:1 | 0.258±0.009ab | 0.307±0.031b | 0.251±0.007a | |
| 5:0 | 0.218±0.014d | 0.328±0.028b | 0.253±0.008a | |
2.3 不同氮素形态配比对甜菜体内氮同化关键酶活性的影响
2.3.1 对NR活性的影响
图2
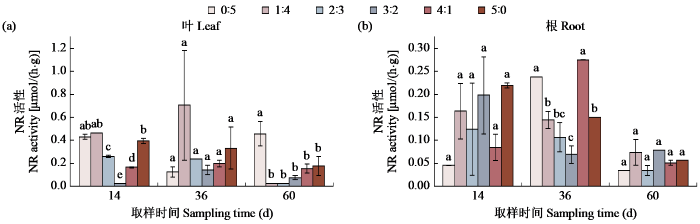
图2
不同氮素形态配比对甜菜NR活性的影响
Fig.2
Effects of different nitrogen form ratio on NR activity of sugar beet
2.3.2 对GDH活性的影响
图3
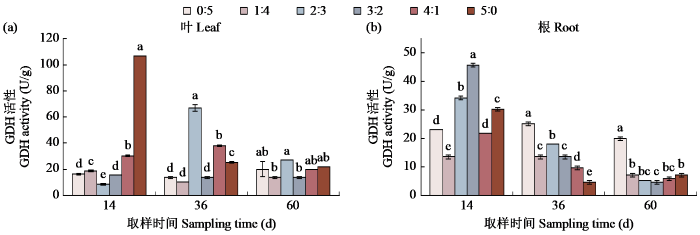
图3
不同氮素形态配比对甜菜GDH活性的影响
Fig.3
Effects of different nitrogen form ratio on GDH activity in sugar beet
2.3.3 对GS活性的影响
图4
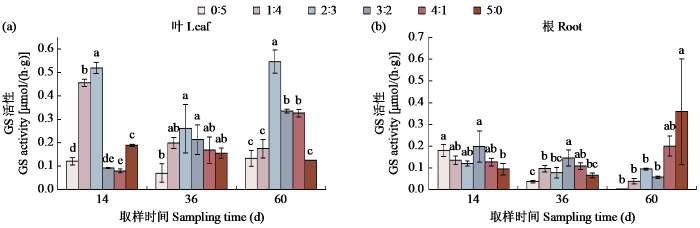
图4
不同氮素形态配比对甜菜GS活性的影响
Fig.4
Effects of different nitrogen form ratio on GS activity of sugar beet
2.4 氮同化关键酶与各项指标之间的相关性分析
根据图5可知,NR与GS活性呈负相关关系,在进行氮同化的过程中,硝酸还原作用和GS作用不是同步进行的,进一步可以说明NR的活性在一定程度抑制了铵同化的进行。除此之外,NR活性与各个表型之间几乎都呈正相关关系,表明NR活性的升高可以明显促进植株的生长。GDH活性与GS活性之间呈负相关关系,表明甜菜在进行铵同化过程中,2种酶存在竞争关系。其中GDH活性与叶绿素含量之间存在正相关关系,表明GS和GDH活性的升高可以明显促进植株体内糖类物质的积累,并对植物光合起到一定促进作用。
图5
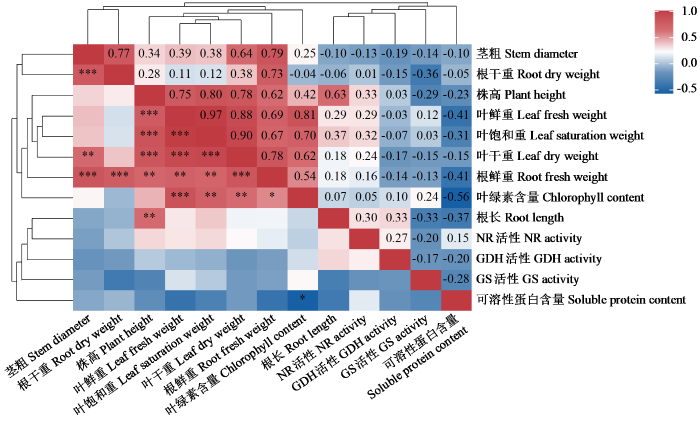
图5
各指标间的相关性分析
红色表示正相关关系,蓝色表示负相关关系;“*”表示P < 0.05显著相关,“**”表示P < 0.01显著相关,“***”表示P < 0.001显著相关。
Fig.5
Correlation analysis among indicators
Red indicates a positive correlation, blue indicates a negative correlation;“*”means a significant correlation in P < 0.05,“**”means a significant correlation in P < 0.01, and“***”means a significant correlation in P < 0.001.
2.5 各指标的主成分分析和隶属函数分析
不同氮素形态下甜菜生长的指标是不同的,不能根据其中的某一指标去判断不同氮素形态对甜菜生长的影响,运用主成分分析和隶属函数综合、客观地得出不同氮素形态对甜菜生长的各项指标影响以及占比情况,判断不同氮素形态与甜菜生长之间的关系,方便进一步探究其内部机制。
由表3可知,通过降维处理将所测得的所有指标分为4个主成分,3个主成分因子可代表全部信息的83.496%,其中,第1主成分代表总方差的42.312%,占比较大的分别为根干重、茎粗、株高、根长、叶饱和重、叶干重等表型指标,表明第1主成分主要反映的是甜菜生长的形态特征。第2主成分代表总方差的16.935%;第3主成分代表总方差的14.768%,这2个主成分中根长、GDH活性、NR活性、叶绿素含量和可溶性蛋白含量的载荷系数较大,表明第2和第3主成分主要反映的是氮同化相关酶的活性以及合成相关物质的积累。
表3 各指标的主成分分析
Table 3
| 指标Index | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| 叶干重Leaf dry weight | 0.929 | -0.128 | -0.005 | 0.257 |
| 叶鲜重Leaf fresh weight | 0.923 | 0.237 | -0.206 | 0.183 |
| 叶饱和重Leaf saturation weight | 0.918 | 0.220 | -0.083 | 0.254 |
| 根鲜重Root fresh weight | 0.883 | -0.345 | -0.013 | -0.191 |
| 株高Plant height | 0.836 | 0.167 | 0.320 | 0.092 |
| 叶绿素含量Chlorophyll content | 0.705 | 0.303 | -0.470 | -0.056 |
| 根干重Root dry weight | 0.428 | -0.752 | 0.262 | -0.299 |
| 茎粗Stem diameter | 0.617 | -0.678 | -0.043 | -0.173 |
| 根长Root length | 0.374 | 0.524 | 0.516 | -0.279 |
| GDH活性GDH activity | -0.035 | 0.515 | 0.348 | -0.463 |
| GS活性GS activity | -0.113 | 0.214 | -0.828 | 0.084 |
| NR活性NR activity | 0.271 | 0.334 | 0.518 | 0.384 |
| 可溶性蛋白含量Soluble protein content | -0.419 | -0.347 | 0.368 | 0.674 |
| 贡献率Contribution rate (%) | 42.312 | 16.935 | 14.768 | 9.481 |
| 累计贡献率 Accumulated contribution rate (%) | 42.312 | 59.247 | 74.015 | 83.496 |
| 因子权重Factors weight | 0.423 | 0.169 | 0.148 | 0.095 |
将6种氮素形态配比下生长甜菜的各项指标进行主成分分析,如图6所示。将6个氮素形态配比分为6个类群,在纯硝态氮供应下生长的甜菜综合指标最好。综合性状最差的是铵硝比为5:0处理下生长的甜菜植株。
图6
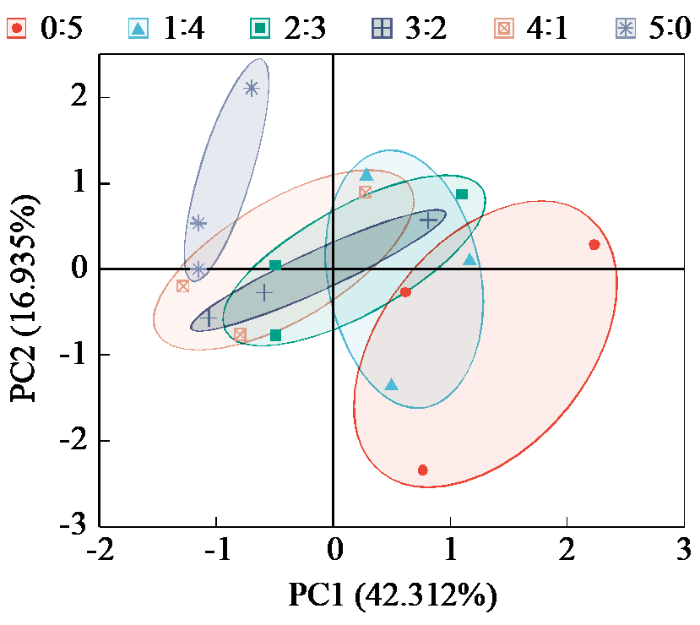
图6
不同氮素形态配比下甜菜各指标的主成分分析
Fig.6
Principal component analysis of various indexes of sugar beet under different nitrogen form ratio
通过隶属函数计算发现,14 d时最高综合得分(D值)为0.737,为纯硝态氮供应下生长的甜菜;36 d时最高D值为0.638,为铵硝比为1:4条件下生长的甜菜;60 d时最高D值是0.500,为铵硝比0:5条件下生长的甜菜。结果表明,在甜菜生长的大部分时期,纯硝态氮的供应可以更好地满足甜菜正常生长的要求,在甜菜旺盛生长时期适量铵态氮更适合甜菜生长。
3 讨论
氮同化的组成部分参与植物生长发育的不同方面。NR是植物体内影响氮素代谢的重要调节酶,催化NO3−生成NO2−,是NO3−同化过程中的第一个酶,也是氮同化中的限速酶,对于植物氮素的代谢具有关键作用[10]。本研究表明,NR和甜菜的表型之间有较大的相关性,NR活性较高,根长、叶饱和重、叶鲜重等表型性状较好。林照辉[11]和刘娜等[12]研究表明,硝态氮的浓度会影响NR活性,其活性随硝态氮的浓度增加而升高,随铵态氮的浓度增加而降低。在甜菜生长旺盛的时期,NR在加入少量铵态氮,及铵硝比为1:4时表现出极高的活性。铵态氮的适量供应促进了铵同化的过程,铵同化的进行使硝酸还原作用的产物减少,在化学反应方向上诱导了NR活性的提高。卢凤刚等[13]研究也同样表明铵态氮的比例升高会降低NR活性。在继续施入铵态氮的过程中,NR活性有着明显的降低。本研究表明,在纯铵态氮供应的条件下,根部NR活性远大于叶部。在此基础上设置不同氮素形态来进一步研究,发现硝态氮与铵态氮对于甜菜不同部位的影响不同,一般来说,外部硝酸盐供应水平较低时,根中硝酸盐的还原比例较大,当外部硝酸盐供应水平较高时,叶部硝酸盐的还原比例较大。在甜菜生长第二个时期,存在少量铵态氮的情况下NR活性高于仅存在硝态氮的情况,这一时期,叶绿素的含量也发生了很大的变化。由此可以得出,在NR活性的研究过程中光照对其存在一定的影响,NR表现出光依赖性,光照强度增加或减少会对NR活性造成一定的影响。光合产物为NO3−提供NADH及还原态铁氧化蛋白。随着铵态氮浓度的增高,不仅对NR的诱导起到一定的抑制作用,对光合作用的抑制也同样导致NR活性的降低。
NR先将植物吸收的硝态氮在植物根或叶肉细胞质中还原成NO2−,然后在质体中将NO2−还原成NH4+,再进一步由GS和谷氨酸合成酶(GOGAT)循环(GS/GOGAT)转化形成有机态谷氨酰胺,然后再形成植物所需要的氨基酸[14]。研究[15]发现,GDH和GS活性的增加可以提高甜菜体内的叶绿素含量,进而影响甜菜光合作用的进行。在铵硝比2:3时,体内的叶绿素含量较多,在铵硝比4:1时甜菜体内的可溶性蛋白含量达到最大。表明铵态氮的施加缩短了无机氮转化为氨基酸的过程,加速了可溶性蛋白的合成。张宏纪等[16]认为,相比硝态氮,铵态氮更有利于增加GS活性,陈煜等[10]认为混合态氮和铵态氮处理大豆叶片的GS活性比硝态氮处理的高,本研究表明在甜菜中只存在硝态氮时GS活性明显低于有铵态氮存在的情况,随着铵态氮比例上升GS活性也随之升高,当铵硝比3:2时其活性达到最高值,可见,硝酸盐的还原和铵同化的同步进行才能促进GS活性提高,硝酸盐代谢还原作用促进了氨化作用的进行。若继续增加铵态氮浓度GS活性反而下降,但在60 d时发现GS活性在铵硝比0:5时达到了最大值,说明GS对氨有着很高的亲和力,随着氨在植物体内的不断累积,迅速将植物组织中的氨同化,避免因为铵态氮过多造成氨毒害。
GDH是谷氨酸合成与代谢过程的关键酶[17],但这一过程会产生NH4+[18],过多的NH4+会对植物的生长产生毒害作用。但也有研究[19]表明GDH可能是GS在遭遇不利情况从而导致活性降低的一种补偿,对GS/GOGAT代谢起协助作用,在14 d时,纯铵态氨处理下GDH的活性最高,即在铵态氮浓度较高的情况下,GDH作为一个具有补偿作用的酶,对整个铵同化的途径起到一定的调控作用,它可以通过催化谷氨酸的合成,增加脯氨酸的数量,进而来提高植物的耐逆性[20]。在铵态氮浓度过高时会通过提升自己的活性来加速铵同化途径的进行,避免产生毒害作用。本试验的相关性分析中可以看出GDH与GS活性之间呈显著的负相关关系,证明在植物的氮同化途径中,两者存在一个竞争关系,但有可能因为过高的GDH活性会产生较多NH4+,所以在大多数氮同化途径中,通常进行的是GS/GOGAT代谢途径。
4 结论
在不同氮素形态配比下,甜菜内部的NR、GS、GDH等3种氮同化关键酶分别通过影响植物表型及相关生理生化指标来调控其生长过程。在纯硝态氮的供应下,甜菜体内3种氮同化关键酶稳定且高效地发挥作用,进而影响相关的其他指标,使得甜菜更好地生长。
参考文献
不同氮素水平对甜菜氮代谢酶和可溶性蛋白含量的影响
DOI:10.11924/j.issn.1000-6850.casb15040052
[本文引用: 1]

为了筛选合适的氮素施用量,研究氮素营养和氮素同化关键酶的关系,以尿素为氮源,采用HI003为试材,研究了不同的施氮水平对甜菜叶片、块根中可溶性蛋白含量、硝酸还原酶和谷氨酰胺合成酶活性的影响。结果表明:叶片中,硝酸还原酶的活性随着施氮水平的提高而增加,但是超过200 kg/hm<sup>2</sup>酶活性下降;块根中硝酸还原酶活性在0~120 kg/hm<sup>2</sup>随着施氮量增加而上升,超过160 kg/hm<sup>2</sup>酶活力下降。叶片、块根中,整个生育期施氮肥处理增加了谷氨酰胺合成酶活性。试验中发现施氮量和生育期早期的叶片中硝酸还原酶和谷氨酰胺合成酶活性呈显著正相关。施氮量和甜菜叶片可溶性蛋白含量在7月15日和9月15日呈显著正相关;在生育期内,氮素水平和块根中的可溶性蛋白含量呈极显著正相关。
Evidence that glutamate acting on presynaptic type-II metabotropic glutamate receptors alone does not fully account for the phenomenon of depolarisation-induced suppression of inhibition in cerebellar Purkinje cells
Understanding regulatory networks and engineering for enhanced drought tolerance in plants
DOI:10.1016/j.pbi.2006.01.019
PMID:16483835
[本文引用: 1]
Drought stress is one of the major limitations to crop productivity. To develop crop plants with enhanced tolerance of drought stress, a basic understanding of physiological, biochemical and gene regulatory networks is essential. Various functional genomics tools have helped to advance our understanding of stress signal perception and transduction, and of the associated molecular regulatory network. These tools have revealed several stress-inducible genes and various transcription factors that regulate the drought-stress-inducible systems. Translational genomics of these candidate genes using model plants provided encouraging results, but the field testing of transgenic crop plants for better performance and yield is still minimal. Better understanding of the specific roles of various metabolites in crop stress tolerance will give rise to a strategy for the metabolic engineering of crop tolerance of drought.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}