


转录因子(transcription factors,TF)能够与启动子上的光响应元件(G-box)等顺式作用元件结合,调控下游基因的表达[4]。近年来,越来越多转录因子的功能被挖掘,其中Dof蛋白是一类植物特有的转录因子,Dof基因家族属于单锌指蛋白超家族,一般由200~400个氨基酸构成[5],因其N末端含有一个C2C2型锌指结构,被称为Dof(DNA binding with one finger)[6]。Dof蛋白由N末端的Dof结构域和C末端的转录调节域构成[7]。由于其C端的氨基酸序列不具备保守性,不同种类的氨基酸能与不同的DNA序列结合,且该区域也是调控基因表达的重要部分,这使得Dof蛋白的功能具有多样性。而N端大约由52个氨基酸组成,又叫保守结构域,能够特异识别序列5′-AAAG-3′的顺式作用元件[8⇓-10]。在该结构域中有4个保守的Cys残基[11]。
Dof转录因子参与植物种子胚乳发育过程中种子储藏蛋白合成、种子萌发、防御机制、碳氮代谢相关基因光调控、叶片组织分化及衰老等过程,在植物的生长发育中起着关键作用[12-13]。植物中发现的第1个Dof基因是ZmMNB1,它通过结合MNF1启动子上的AAGG序列进而参与C4植物光合作用的表达调控[14]。此后,越来越多的植物Dof基因被深入研究。如过表达OsDof 12基因可以使转基因水稻的花期提前,在长日照条件下提前开花[15],过表达GhDof 1基因能够上调与胁迫相关基因GhSOD及GhMYB的表达量,从而提高棉花的耐低温和耐盐胁迫的能力[16],番茄SlDof 22与SlSOS1基因的启动区序列结合,参与抗坏血酸积累和盐胁迫反应[17]。拟南芥OBP1调控植物防御基因的表达[18]。11个ZmDof基因的表达模式都受到盐或干旱胁迫的诱导或抑制[19],表明这些基因广泛参与干旱胁迫的响应途径。关于Dof基因的克隆及功能研究主要集中在拟南芥、番茄、水稻和烟草中[20],高粱中有关Dof基因的功能以及该基因家族成员的挖掘鲜见报道。
本研究利用生物信息学技术挖掘高粱Dof基因家族成员,并对其染色体定位、基因结构、理化性质、进化关系和启动子元件等进行分析,并预测了蛋白互作关系,为进一步探究Dof家族成员的功能和响应干旱作用机制提供参考,为培育新品种提供分子资源。
1 材料与方法
1.1 试验材料
试验材料为河南省农业科学院粮食作物研究所培育的高粱新品种豫粱10号。干旱胁迫试验:高粱种子经75%酒精及5% NaClO消毒处理10 min,无菌水冲洗后置于培养箱中萌发,每日设置光照16 h、黑暗8 h,温度25 ℃。将发芽一致的种子移到Hoagland营养液中进行后培养,每3 d更换1次营养液,将生长2周且长势一致的高粱幼苗用20% PEG6000溶液处理,分别在24、48、72 h后取高粱的叶片,将其放入液氮中冷冻,于-80 ℃冰箱保存备用。每个取样时间点设置3个重复。
1.2 RNA提取与定量PCR
按照RNA试剂盒方法提取高粱RNA,用NanoDrop2000检测RNA浓度与纯度,用10 g/L琼脂糖胶验证RNA提取的完整度。通过反转录试剂盒将提取的RNA反转成cDNA,荧光定量PCR检测SbDof基因的表达。荧光定量试剂盒采用abm SYBR Green Realtime PCR Master Mix,反应体系总体积20.0 μL,含BlasTaqTM Mix(2×)10.0 μL、上游和下游引物(10 μmol/L)各0.5 μL、cDNA模板1.0 μL、无菌蒸馏水8.0 μL。反应条件:95 ℃预变性2 min;95 ℃变性15 s,60 ℃退火40 s,循环40次;每个样品均3次重复。计算公式为∆Ct目标基因=Ct目标基因-Ct同一样本Actin;∆∆Ct目标基因=处理组∆Ct目标基因-对照组∆Ct目标基因。基因相对表达量=2-∆∆Ct。Actin作为内参基因。表1为差异表达引物的序列。
表1 高粱Dof基因的qRT-PCR扩增所用引物序列
Table 1
| 基因Gene | 登录号Accession number | 上游引物序列Upstream primer sequence | 下游引物序列Downstream primer sequence |
|---|---|---|---|
| SbDof-1 | Sobic.001G034300 | GTCGTCCAAGTCAAACTCGTC | TCAGTCCGGGGAACCCTAA |
| SbDof-4 | Sobic.001G179300 | CGACTACCACCACCAGCAG | CAGCTTCCAGTGCAGATCCT |
| SbDof-8 | Sobic.001G489900 | CTCATCAAGCTGTTCGGCAAG | GTCTTCAGTGTCCGCAACCT |
| SbDof-10 | Sobic.002G313800 | TCCATGAACCCTGCGGTTAG | TAGAAGAGCGTGGCAATGGA |
| SbDof-11 | Sobic.002G421900 | GAGCTGCGTCCCCTCG | ACCCTCCGTCTTGGCTATCT |
| SbDof-13 | Sobic.003G121400 | CCTGGGATCAAGCTCTTCGG | GGCGCATCGACCTTCATTTC |
| SbDof-24 | Sobic.007G223200 | ACAACTACAACACCTCCCAGC | GTGTCTGTGACACCACTGGC |
| SbDof-25 | Sobic.008G001700 | TCTCAGGCTCTGGTTCCTCT | CGGGAGCACATTGGGGAATA |
| Actin | LOC110436378 | TGAAGTGCGACGTGGATATTAGGA | GCTGGAATGTGCTGAGAGATGC |
1.3 高粱Dof基因家族成员基因序列分析
在PlantTFDB数据库(
1.4 高粱Dof基因的二级结构
利用SOPMA(
1.5 高粱Dof基因启动子分析
从NCBI数据库中提取每个Dof基因转录起始位点(TSS)上游区域的2000 bp,上传到PlantCARE数据库中,利用PlantCARE数据库预测Dof基因的启动子区顺式元件,并对启动子的数量进行统计,使用TBtools绘图分析该家族成员的顺式作用元件。
1.6 高粱Dof基因系统进化与基因结构分析
通过MEGA11中的Clustal X对高粱Dof基因家族的蛋白序列进行比对,利用Clustal X2对高粱、水稻、拟南芥3个物种所有的Dof蛋白序列进行多重序列比对,利用Neighbor-Joining(邻接法)构建系统进化树。并利用GSDS(
1.7 高粱Dof基因保守基序分析
利用在线保守基序分析软件MEME(
1.8 高粱Dof基因的组织表达
利用Expression Atlas(
1.9 Dof家族基因的互作蛋白预测
利用在线工具STRING(
2 结果与分析
2.1 高粱Dof基因基本信息及染色体定位
根据PlantTFDB获取的高粱基因组信息,通过结构域验证最终获取了30个SbDof基因,通过Tbtools软件分析30个Dof基因在染色体上的分布情况。结果(图1)表明,高粱的30个Dof基因分布在高粱的9条染色体上,每条染色体上基因数量为1~8个。其中Chr01上基因数目最多(8个),其次是Chr03(6个);Chr02、Chr04、Chr08、Chr09上均含有3个基因;Chr06上含有2个基因,而Chr05、Chr07上均仅含有1个基因。根据该家族成员在染色体位置分别命名为SbDof1~SbDof30,同时对SbDof家族成员信息进行分析,结果如表2所示,SbDof家族成员的基因长度差异也是较大的,范围在923~3309 bp,SbDof基因编码的氨基酸长度在168(SbDof15)~560(SbDof13)之间,氨基酸序列的平均长度为349;蛋白质等电点介于4.75(SbDof17)和10.27(SbDof15)之间,其中SbDof6、SbDof11、SbDof13、SbDof17、SbDof24和SbDof29等电点小于7.0,呈酸性;其余24个SbDof蛋白等电点大于7.0,呈碱性,蛋白质的相对分子质量为17 251.49~61 403.46 Da,平均值约为36.18 kDa。TMPred和TMHMM在线软件预测的结果一致,表明高粱Dof蛋白均不存在跨膜区,即都是非跨膜蛋白。30个编码蛋白基因中均不含信号肽区域,这表明高粱Dof蛋白不属于分泌蛋白。亚细胞分析结果表明,30个SbDof基因都位于细胞核中。
图1
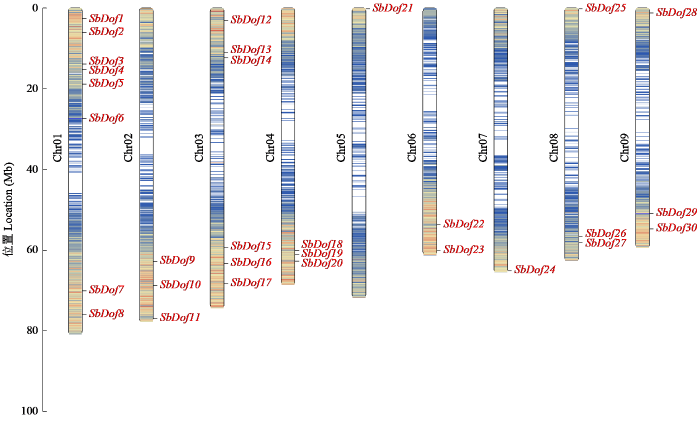
图1
高粱Dof家族基因在染色体上的位置
Fig.1
Location of sorghum Dof family genes on chromosomes
表2 高粱Dof基因家族成员基本信息
Table 2
| 基因编号 Gene code | 基因ID Gene ID | 氨基酸数 Number of amino acids | 基因长度 Gene length (bp) | 编码区 Coding region | 等电点 Isoelectric point | 蛋白质相对分子质量 Protein relative molecular mass (Da) | 信号肽分析 Signal peptide analysis |
|---|---|---|---|---|---|---|---|
| SbDof1 | Sobic.001G034300 | 435 | 3309 | 1572~2879 | 9.34 | 43 307.61 | 0.0031 |
| SbDof2 | Sobic.001G078700 | 380 | 1995 | 365~1507 | 9.23 | 38 671.80 | 0.0008 |
| SbDof3 | Sobic.001G166200 | 351 | 1397 | 94~1149 | 8.93 | 35 519.91 | 0.0025 |
| SbDof4 | Sobic.001G179300 | 385 | 1950 | 419~1576 | 8.34 | 38 461.74 | 0.0143 |
| SbDof5 | Sobic.001G206300 | 261 | 1535 | 484~1269 | 9.32 | 26 378.44 | 0.0026 |
| SbDof6 | Sobic.001G252800 | 476 | 2136 | 315~1745 | 6.31 | 49 778.02 | 0.0013 |
| SbDof7 | Sobic.001G420300 | 394 | 1997 | 298~1485 | 9.94 | 40 285.77 | 0.0015 |
| SbDof8 | Sobic.001G489900 | 424 | 1810 | 259~1533 | 8.65 | 45 170.80 | 0.0016 |
| SbDof9 | Sobic.002G236600 | 252 | 1348 | 127~1077 | 9.28 | 25 792.75 | 0.0109 |
| SbDof10 | Sobic.002G313800 | 354 | 2489 | 547~1611 | 8.16 | 34 828.74 | 0.0137 |
| SbDof11 | Sobic.002G421900 | 484 | 1980 | 137~1591 | 6.35 | 51 627.69 | 0.0020 |
| SbDof12 | Sobic.003G033500 | 235 | 1519 | 198~1142 | 9.90 | 24 113.61 | 0.0619 |
| SbDof13 | Sobic.003G121400 | 560 | 2222 | 208~1890 | 5.14 | 61 403.46 | 0.0005 |
| SbDof14 | Sobic.003G131100 | 512 | 2359 | 347~1885 | 9.04 | 54 771.55 | 0.0025 |
| SbDof15 | Sobic.003G253200 | 168 | 2049 | 1425~1940 | 10.27 | 17 251.49 | 0.0015 |
| SbDof16 | Sobic.003G301700 | 260 | 1645 | 471~1253 | 9.21 | 27 855.15 | 0.0011 |
| SbDof17 | Sobic.003G367100 | 335 | 1495 | 218~1225 | 4.75 | 35 258.15 | 0.0260 |
| SbDof18 | Sobic.004G254000 | 304 | 1476 | 235~1149 | 8.81 | 30 986.43 | 0.0212 |
| SbDof19 | Sobic.004G266200 | 295 | 1639 | 444~1331 | 8.51 | 30 934.23 | 0.0020 |
| SbDof20 | Sobic.004G284400 | 457 | 2244 | 460~1833 | 9.28 | 45 947.04 | 0.0002 |
| SbDof21 | Sobic.005G001500 | 340 | 1527 | 281~1303 | 9.11 | 36 180.21 | 0.0011 |
| SbDof22 | Sobic.006G182300 | 388 | 2534 | 696~1811 | 8.76 | 38 945.94 | 0.0013 |
| SbDof23 | Sobic.006G267900 | 224 | 1682 | 702~1376 | 7.59 | 23 265.65 | 0.0374 |
| SbDof24 | Sobic.007G223200 | 265 | 1502 | 161~1020 | 5.86 | 27 579.64 | 0.0062 |
| SbDof25 | Sobic.008G001700 | 354 | 1524 | 234~1298 | 9.12 | 38 038.01 | 0.0020 |
| SbDof26 | Sobic.008G136100 | 340 | 2016 | 610~1632 | 9.05 | 34 557.65 | 0.0100 |
| SbDof27 | Sobic.008G148101 | 275 | 923 | 96~923 | 9.77 | 28 033.91 | 0.0214 |
| SbDof28 | Sobic.009G014400 | 385 | 2066 | 428~1585 | 9.07 | 39 250.54 | 0.0028 |
| SbDof29 | Sobic.009G153400 | 366 | 1472 | 117~1217 | 5.68 | 38 488.27 | 0.0010 |
| SbDof30 | Sobic.009G196100 | 216 | 1909 | 419~1087 | 9.99 | 22 783.69 | 0.0014 |
2.2 高粱Dof家族蛋白二级结构分析
高粱Dof家族成员的二级结构主要由α-螺旋和无规则卷曲构成,其中无规则卷曲占比最高,而延伸链(延伸链是β-折叠的组成结构)、β-转角占比较低(表3)。由此推测,高粱Dof蛋白的主要结构元件是α-螺旋和无规则卷曲,而延伸链和β-转角是分散于蛋白中的。
表3 高粱Dof蛋白二级结构分析
Table 3
| 基因号 Gene ID | α-螺旋 α-helix (Hh) | 延伸链 Extended strand (Ee) | β-转角 β-turn (Tt) | 无规则卷曲 Random coil (Cc) | 基因号 Gene ID | α-螺旋 α-helix (Hh) | 延伸链 Extended trand (Ee) | β-转角 β-turn (Tt) | 无规则卷曲 Random coil (Cc) |
|---|---|---|---|---|---|---|---|---|---|
| SbDof1 | 12.87 | 8.97 | 4.37 | 73.79 | SbDof16 | 16.92 | 14.23 | 5.77 | 63.08 |
| SbDof2 | 26.05 | 10.53 | 5.26 | 58.16 | SbDof17 | 19.10 | 8.96 | 4.48 | 67.46 |
| SbDof3 | 23.36 | 13.96 | 7.12 | 55.56 | SbDof18 | 22.37 | 15.13 | 4.93 | 57.57 |
| SbDof4 | 33.25 | 11.43 | 4.94 | 50.39 | SbDof19 | 24.75 | 12.88 | 5.08 | 57.29 |
| SbDof5 | 26.05 | 11.88 | 5.75 | 56.32 | SbDof20 | 17.94 | 15.10 | 6.13 | 60.83 |
| SbDof6 | 17.23 | 9.24 | 3.15 | 70.38 | SbDof21 | 15.88 | 14.41 | 4.12 | 65.59 |
| SbDof7 | 17.01 | 12.69 | 5.58 | 64.72 | SbDof22 | 11.34 | 11.60 | 2.84 | 74.23 |
| SbDof8 | 13.68 | 8.02 | 1.65 | 76.65 | SbDof23 | 34.38 | 8.48 | 8.04 | 49.11 |
| SbDof9 | 11.90 | 17.46 | 6.75 | 63.89 | SbDof24 | 25.28 | 14.34 | 10.19 | 50.19 |
| SbDof10 | 36.72 | 9.60 | 6.50 | 47.18 | SbDof25 | 14.41 | 15.54 | 3.95 | 66.10 |
| SbDof11 | 13.84 | 7.23 | 1.45 | 77.48 | SbDof26 | 26.47 | 11.76 | 7.94 | 53.82 |
| SbDof12 | 13.62 | 15.32 | 10.21 | 60.85 | SbDof27 | 27.64 | 11.64 | 7.27 | 53.45 |
| SbDof13 | 13.75 | 6.07 | 2.14 | 78.04 | SbDof28 | 24.16 | 12.47 | 10.91 | 52.47 |
| SbDof14 | 19.73 | 8.20 | 3.12 | 68.95 | SbDof29 | 26.23 | 12.84 | 8.47 | 52.46 |
| SbDof15 | 30.36 | 6.55 | 4.76 | 58.33 | SbDof30 | 27.78 | 16.20 | 5.56 | 50.46 |
2.3 高粱Dof基因系统进化与基因结构分析
将鉴定到的高粱Dof基因家族成员进行氨基酸序列分析(图2),根据已知拟南芥Dof基因家族成员分类信息对高粱Dof基因家族进行分类,将其分为5组(І、Ⅱ、Ⅲ、Ⅳ、V),其中І组成员最多,包含12个家族成员,Ⅱ、Ⅲ、Ⅳ、V组分别包含3、5、8、2个,同组内Dof成员之间蛋白序列一致性较高。分析了高粱30个Dof家族成员的结构,SbDof大部分的基因家族成员含有1~2个内含子(图3)。将鉴定的10个保守基序命名为Motif1~Motif10(图4)。在所有的Dof家族成员中都包含Motif1结构,即有CX2CX21CX2C保守基序(图5),表明这个基序可能为Dof的核心保守结构域,且该结构域在进化过程中较为稳定,对Motif1的保守基序序列分析结果表明该序列为锌指结构。除Motif1外,І组中有5个Dof家族成员均含有Motif2,有7个Dof家族成员均含有Motif3,IV组中SbDof28、SbDof7、SbDof1均含有Motif8,同一亚组高粱的Dof成员有类似Motif组成特征。在相同组中含有相同保守基序的Dof蛋白也许具有相似的功能。
图2
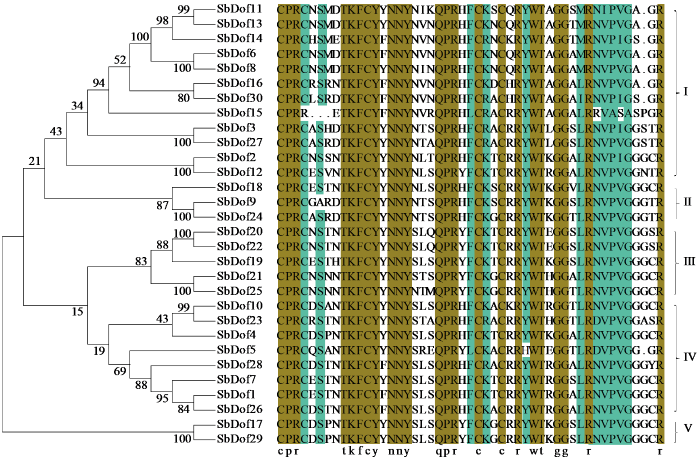
图2
SbDof蛋白家族保守序列比对分析
Fig.2
Comparison and analysis of conserved sequences of SbDof protein family
图3
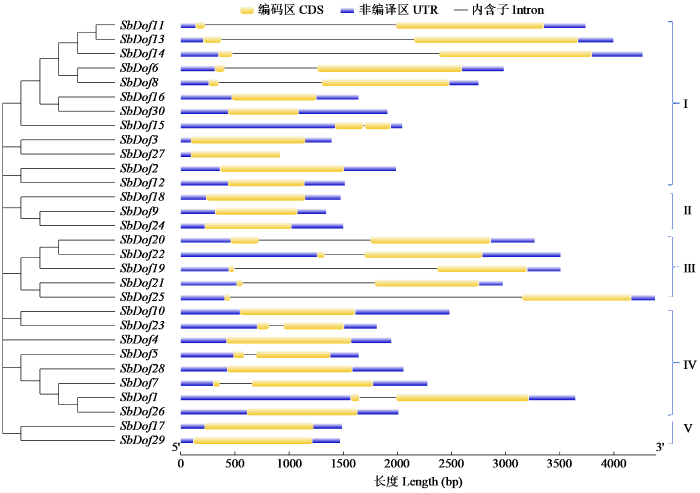
图4

图4
高粱Dof家族蛋白保守基序分析
Fig.4
Analysis of conserved motifs of sorghum Dof family proteins
图5

图5
高粱Dof基因的功能域序列分析
Fig.5
Sequence analysis of functional domain of Dof gene in sorghum
2.4 高粱Dof基因启动子分析
如图6所示,该基因家族成员启动子区域含有许多顺式作用元件,这些元件与其他蛋白结合并激活各种途径,除了常见的光响应元件,还有与植物激素及非生物胁迫相关的顺式作用元件。鉴定到的胁迫相关顺式元件主要包括ABRE(脱落酸响应元件)、ARE(厌氧诱导响应元件)、CAT-box(分生组织表达)、CGTCA-motif(茉莉酸甲酯响应元件)、G-box(光响应元件)、RY-element(种子特异调控元件)、TGA-box(生长素响应元件)、MBS(干旱诱导响应元件)、LTR(低温响应元件)和circadian(参与昼夜节律控制的顺式调控元件)。这表明该家族成员可能参与非生物胁迫和植物激素的严格调控,在高粱的生长发育中发挥着重要作用。
图6
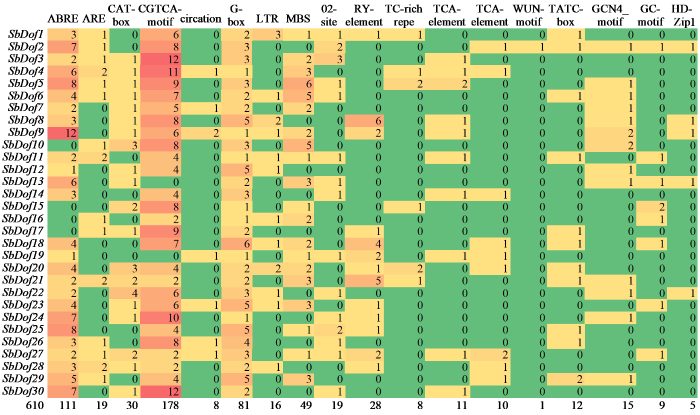
图6
高粱Dof家族基因顺式调控元件分析
Fig.6
Analysis of cis-regulatory elements of Dof family genes in sorghum
30个SbDof家族基因成员中共检测到610个主要顺式调控元件,CGTCA-motif和ABRE在激素反应元件中所占比例较大,CGTCA-motif数量最多,共检测到178个;其次为ABRE,检测到111个。因此,SbDof基因可能对CGTCA-motif和脱落酸激素更敏感。在组织特异性顺式作用元件中,种子特异性表达元件和分生组织表达元件所占比例较高,其中RY-element共检测到28个,CAT-box共检测到30个,说明部分SbDof基因家族成员可能在高粱种子发育或细胞的组织分化过程中发挥作用。G-box数量共检测到81个,表明该基因受光调控影响也较大。在非生物胁迫响应元件中,LTR共检测到16个,MBS共检测到49个,这表明一些SbDof基因家族成员可能在植物适应各种胁迫中发挥作用。
2.5 高粱Dof基因系统进化树
利用MEGA软件对30个高粱Dof(SbDof1- 30)、44个玉米Dof(ZmV3Dof1-44)及30个拟南芥Dof(AtDof1.1-5.7)基因家族成员进行进化树构建。如图7所示,以拟南芥Dof家族基因为参考,将高粱Dof基因分为了6组,I组内包括高粱Dof成员20个,拟南芥Dof成员31个,玉米Dof成员28个;II和III组均包括2个高粱Dof成员和3个玉米成员;V组有1个高粱成员、2个玉米成员;VI组有1个高粱成员和1个玉米成员,II、III、V、VI组内均不包括拟南芥的家族成员,表明这4个分支的Dof蛋白也许是在玉米、拟南芥及高粱共同祖先的进化过程中,在玉米及高粱中保存下来,而在拟南芥中丢失了。在IV组内包括4个高粱Dof成员、9个玉米Dof成员和5个拟南芥Dof成员;除I组中SbDof18和拟南芥AtDof5.4亲缘关系较近外,其他拟南芥Dof家族成员多单独聚为一类,表明高粱Dof家族基因与单子叶植物玉米Dof的亲缘关系较近,与双子叶植物拟南芥Dof基因的亲缘关系较远。
图7
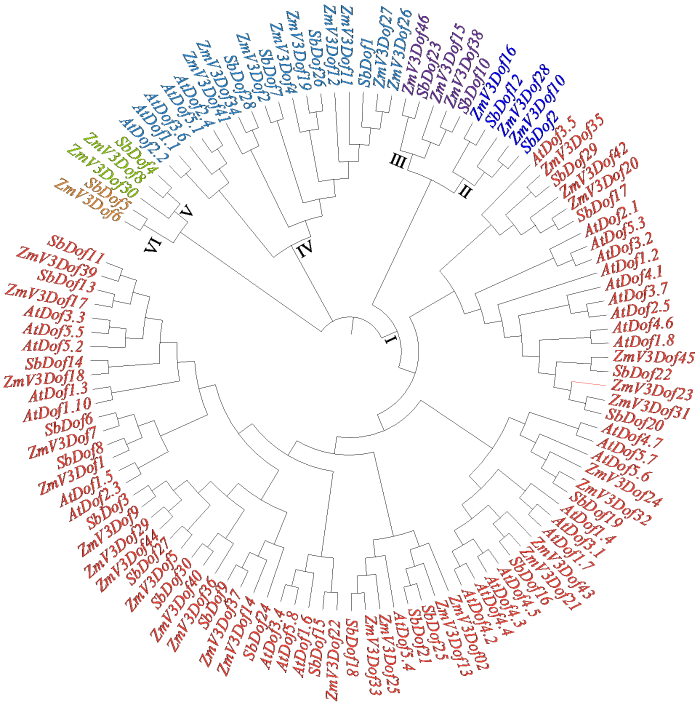
图7
3个作物的Dof基因家族系统进化树
Fig.7
Phylogenetic tree of Dof gene family in three crops
2.6 高粱Dof基因的组织表达分析
高粱Dof基因家族成员组织表达分析结果(图8)表明,SbDof家族成员在高粱不同部位的表达量不同,其中SbDof3、SbDof12、SbDof17、SbDof21、SbDof25、SbDof27、SbDof29和SbDof30 8个基因在叶片中无表达,而位于亚组I中的SbDof8和SbDof11在高粱叶片中的表达量均较高,表明它们可能在高粱叶片的生长发育中起重要作用,可作为高粱生长发育相关的候选基因进一步验证。SbDof24基因在高粱的雌蕊及授粉后第5天播种中的表达量最高,表明这些基因可能与雌配子的发育相关,而SbDof4、SbDof12、SbDof17、SbDof27和SbDof29在高粱的胚乳中均不表达。此外,个别Dof基因的表达具有组织特性,如SbDof21基因仅在胚胎及胚乳中表达。SbDof24基因在高粱雌蕊及授粉后第5天的种子中表达量最高,表明该基因可能在高粱授粉及雌配子的发育中发挥重要作用。SbDof12基因在高粱的早期花序、胚乳、叶片、雌蕊、花药组织中表达量较少或无表达,表明该基因并不参与高粱这些组织的生长发育。其余部分家族成员在各组织部位高表达或低表达,表明该家族在高粱中具有较强的组织表达特异性,从而在高粱的生长发育中发挥不同的作用。
图8
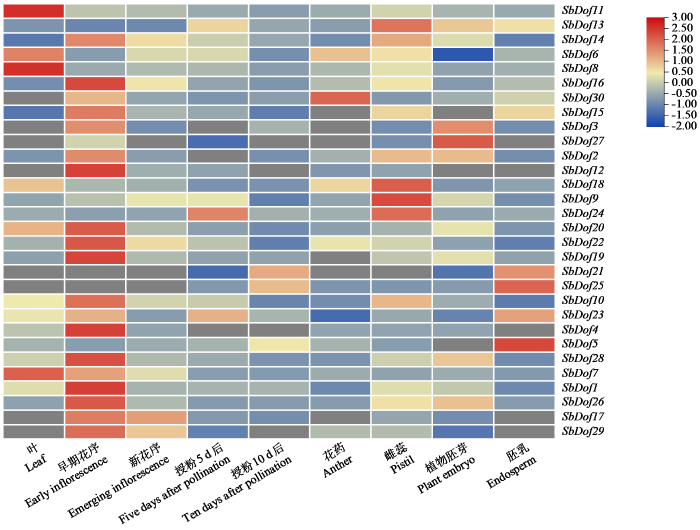
图8
高粱Dof基因家族成员的表达模式分析
Fig.8
The expression patterns of the Dof genes in sorghum
2.7 预测Dof10基因的互作蛋白
为了探究SbDof10基因编码蛋白的互作关系,对SbDof10的互作蛋白进行预测,结果(表4)表明,SbDof10可能与Sb03g034780.1(发病机制相关基因转录激活因子)、Sb01g031890.1(锌指蛋白ZAT10)及Sb09g027800.1(转录因子MYB)等存在互作关系。
表4 SbDof10功能互作蛋白预测
Table 4
| 互作蛋白Interaction protein | 基因描述Gene description | 互作系数Interaction coefficient | 氨基酸数量Number of amino acids |
|---|---|---|---|
| Sb09g006196.1 | Loc8065916亚型x1,响应低温胁迫 | 0.695 | 384 |
| Sb01g029000.1 | WRKY DNA结合域超家族蛋白 | 0.694 | 346 |
| Sb03g034780.1 | 发病机制相关基因转录激活因子PTI5 | 0.663 | 178 |
| Sb01g031890.1 | 锌指蛋白ZAT10 | 0.622 | 253 |
| Sb09g027800.1 | 转录因子MYB | 0.663 | 289 |
2.8 干旱胁迫下SbDof基因家族的表达分析
研究[17]表明,Dof转录因子参与非生物胁迫响应,并在调控逆境相关基因的表达中起重要作用。为进一步研究SbDof基因响应干旱胁迫的表达谱,采用PEG6000模拟干旱胁迫于不同时间点取样,通过qRT-PCR分析SbDof基因处理与对照之间的相对表达值。结果(图9)表明,在30个Dof家族成员中获得了8个差异表达较大的基因,各基因在干旱胁迫处理后不同时间点与对照0 h归一化相比,Dof1基因在干旱处理24及48 h后表达量下调,在72 h后表达量上调,而Dof8在24 h时表达量最高,Dof10基因在24 h表达量下降,在48及72 h时表达量上调极显著,Dof13基因在48 h表达量下调显著,Dof25基因在24及72 h时表达量上调显著,在48 h时下调,而Dof4在72 h时表达量最低,Dof24基因在24 h表达量下调,而在48及72 h表达量上调。这表明不同基因对于不同时间的干旱响应程度不同。
图9
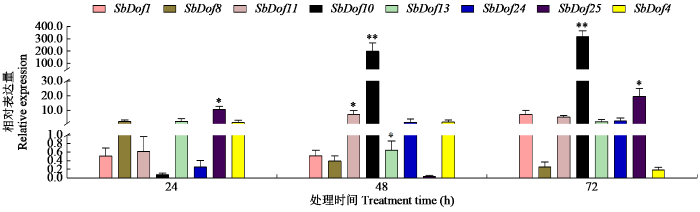
图9
高粱Dof家族基因在干旱处理下的表达分析
“*”表示差异显著(P < 0.05),“**”表示差异极显著(P < 0.01)。
Fig.9
Expression analysis of Dof family genes in sorghum under drought treatments
“*”indicates significant difference (P < 0.05),“**”indicates extremely significant difference (P < 0.01).
3 讨论
本研究利用生物信息学的方法,从高粱基因组中鉴定了30个Dof成员,其中29个SbDof成员含有CX2CX21CX2C保守序列,SbDof15基因保守序列为CX20CX2C。30个SbDof成员不均匀地分布在9条染色体上,第10染色体无分布。氨基酸长度、蛋白质分子质量、等电点等存在差异。
对3个作物的Dof蛋白进行系统进化树分析,将Dof蛋白分为6个亚家族。目前,已有许多关于玉米、拟南芥Dof基因的报道[21],I组中的拟南芥AtDof5.8、AtDof2.3响应盐、干旱等非生物胁迫[22-23],AtDof2.3在PEG胁迫处理后,其表达量是对照的69.46倍,且过表达该基因,经过干旱处理,其根长、过氧化物酶和超氧化物歧化酶等指标较对照显著增加[24];而过表达AtDof5.8增强了转基因拟南芥对干旱及盐胁迫的抗性[25]。同样位于该组的玉米ZmDof41、ZmDOf42、ZmDof7基因参与玉米应答干旱胁迫的响应,但表达模式存在差异,ZmDof7、ZmDOf42在干旱胁迫处理早期没有明显变化,24 h显著诱导上调表达;ZmDof41在干旱处理1 h时受到强烈诱导,显著上调表达,其表达水平为对照的10倍[26]。本研究发现,位于同一分支的SbDof13、SbDof8、SbDof25、SbDof11基因受到干旱胁迫诱导,其中SbDof25基因在24 h的表达量是对照的10倍以上,说明可依据水稻及拟南芥的Dof家族成员相关的功能研究,寻找其同源基因间的关系,为研究高粱Dof家族成员的功能提供基础。
启动子的顺式作用元件分析发现,SbDof家族成员不仅参与光、低温、干旱等非生物胁迫下的转录调控,同时也参与了生长素、茉莉酸甲酯、脱落酸、赤霉素等激素信号传导调控,这些指标都是与胁迫密切相关的响应元件。几乎所有的家族成员都含有MeJA及ABA的响应元件,不同的家族成员可能通过ABA、MeJA等响应元件参与高粱的各种胁迫响应,SbDof基因如何通过这些响应元件参与表达还需要进一步研究。张凤娇[24]研究发现,AtDof2.1能够与AtDof2.3基因启动子上的ABA响应元件结合,从而参与拟南芥的胁迫响应。
基因的组织表达模式往往能为基因功能的研究提供重要信息[27],葛敏等[28]研究发现,ZmDof39基因在玉米叶片中的表达量最高,在玉米的生长发育及其组织分化过程中发挥着重要作用,本研究也发现SbDof11、SbDof8基因在高粱叶片中的表达量最高,并且与ZmDof39基因位于同一分支,推测这2个基因可能与ZmDof39有类似的功能。44个玉米ZmDof基因中,ZmDof3和PBF(ZmDof13)只在玉米籽粒的胚乳中特异表达,在其他组织部位均无表达[29],本研究也发现与ZmDof3和PBF(ZmDof13)位于同一分支的SbDof25及SbDof21基因在高粱的胚乳中表达量较高,由此推测这些Dof基因很可能有相似的功能。
研究[30]发现,Dof基因在植物响应非生物胁迫反应中发挥重要的调控作用,唐跃辉等[31]研究发现,小桐子JcDof3、JcDof5、JcDof17、JcDof20基因在干旱胁迫处理后上调表达,本研究也发现高粱SbDof1、SbDof10、SbDof25基因随干旱处理时间增加,表达量显著上调。SbDof10基因在干旱处理后显著上调表达,在干旱胁迫48 h时SbDof10基因的表达量显著升高,60 h时达到峰值,是对照近260倍。对SbDof10编码的蛋白进行互作蛋白预测,发现与SbDof10具有互作关系的转录因子SbWRKY30通过调控干旱胁迫应答功能基因SbRD19的表达,增强高粱抵抗干旱胁迫的能力[32]。番茄SlPti5通过调节活性氧系统和激素通路,在植物抗灰葡萄孢免疫应答中发挥调控作用[33],本研究也发现SbDof10基因可能与发病机制相关基因转录激活因子PTI5存在互作关系。推测SbDof10可能与这些潜在的互作蛋白有协同作用,表明该基因可能在响应干旱胁迫反应中具有重要的调控作用,这为后期进一步研究SbDof10的功能提供参考依据。
4 结论
本研究在基因组水平上鉴定了30个SbDof高粱转录因子基因家族,并对其基因结构、系统进化、理化性质、结构域、组织表达等方面进行了分析。SbDof家族成员的理化性质存在一定的差异,24个成员的等电点大于7.0,为碱性氨基酸。所有的SbDof都具有保守的C2C2单锌指结构域。SbDof转录因子中锌指结构的大部分氨基酸高度保守。在干旱胁迫条件下该家族成员的表达模式分析发现SbDof10基因显著上调表达,可能与高粱耐旱性有关。这为进一步分析SbDof的功能提供了理论基础,为高粱遗传改良和抗旱机制解析提供新思路。
参考文献
高粱转录因子SbWRKY71基因的克隆及其在逆境胁迫下的表达分析
DOI:10.11869/j.issn.100-8551.2021.07.1532
[本文引用: 1]

为了探究WRKY转录因子在植物抵抗逆境胁迫方面的重要作用,本研究通过基因克隆的方法,从高粱BTx623中克隆得到一个WRKY转录因子基因(SbWRKY71),该转录因子基因全长1 335 bp(phytozome登录号:Sb04g005520),编码364个氨基酸,分子量为38.95 kDa;预测该转录因子定位于细胞核,具有WRKY转录因子典型的保守结构域,且该蛋白属于WRKY蛋白家族的第Ⅱ组成员。系统进化树分析表明,高粱SbWRKY71氨基酸序列与禾本科作物玉米ZmWRKY71的亲缘关系最近,为75%;实时荧光定量PCR(RT-qPCR)检测表明,SbWRKY71基因表达具有组织特异性,在叶中表达丰度最高,茎中最低。经激素水杨酸(SA,1 mmol·L<sup>-1</sup>)、吲哚-3-乙酸(IAA,10 μmol·L<sup>-1</sup>)和脱落酸(ABA,200 μmol·L<sup>-1</sup>)处理后,SbWRKY71的表达量呈现先下降后升高再下降的趋势;在γ-氨基丁酸(GABA)和甘露醇(D-Mannitol,300 mmol·L<sup>-1</sup>)模拟干旱胁迫以及氯化钠(NaCl,250 mmol·L<sup>-1</sup>)盐胁迫处理下,SbWRKY71的表达量均先升后降,分别在3、6和9 h达到最大值;高粱经病原相关分子模式(PAMPs)flg22(100 nmol·L<sup>-1</sup>)、翻译延长因子(elf18,100 nmol·L<sup>-1</sup>)处理后,SbWRKY71表达均受到抑制,但在几丁质(Chitin,8 nmol·L<sup>-1</sup>)处理下,SbWRKY71受到诱导表达。本研究为进一步探索SbWRKY71基因在调节高粱抗性、响应激素以及逆境胁迫应答等过程中的作用机制提供了基础。
Systematic analysis of the bZIP family in tobacco and functional characterization of NtbZIP 62 involvement in salt stress
Diversity and similarity among recognition sequences of Dof transcription factors
A putative flowering- time-related Dof transcription factor gene, JcDof3, is controlled by the circadian clock in Jatropha curcas
Dof domain proteins: plant-specific transcription factors associated with diverse phenomena unique to plants
DOI:10.1093/pcp/pch055
PMID:15111712
[本文引用: 1]
Dof (DNA-binding with one finger) domain proteins are plant-specific transcription factors with a highly conserved DNA-binding domain, which presumably includes a single C(2)-C(2) zinc finger. During the past decade, numerous Dof domain proteins have been identified in both monocots and dicots including maize, barley, wheat, rice, tobacco, Arabidopsis, pumpkin, potato, and pea. Biochemical, molecular biological and molecular genetic analyses revealed that Dof domain proteins function as a transcriptional activator or a repressor involved in diverse plant-specific biological processes. Although more physiological roles of Dof domain proteins would be elucidated in future because of numerous Dof domain proteins in plants, it is already evident that the Dof domain proteins play critical roles as transcriptional regulators in plant growth and development. Here I summarize our current knowledge about Dof domain proteins.
Transcriptome analysis identifies a zinc finger protein regulating starch degradation in kiwifruit
Rice DNA-binding one zinc finger 24 (OsDOF24) delays leaf senescence in a jasmonate-mediated pathway
Dof transcription factors in carrot: genome-wide analysis and their response to abiotic stress
DOI:10.1007/s10529-015-1966-2
PMID:26466595
[本文引用: 1]
The DNA-binding one zinc finger (Dof) family transcription factors (TF) are involved in stress response. Dof TFs in carrot were identified and the responses of DcDof genes to abiotic stresses were analyzed.46 DcDofs in carrot were identified from carrot genome database. Based on the conserved domain in Dof TF family of Arabidopsis thaliana, the DcDof TFs were divided into four classes, named class A, B, C and D. Carrot and Arabidopsis shared most motifs in the same subgroup. Real-time quantification PCR analysis showed tissue-specific expression patterns in DcDofs. DcDofs from eight subgroups responded to four abiotic stress treatments.The expression profiles were different with the abiotic stresses changed, indicating complicated regulatory mechanisms in Dof TF family in higher plant, and the response mechanisms of Dof genes may be influenced by different plant species.
Overexpression of GhDof1 improved salt and cold tolerance and seed oil content in Gossypium hirsutum
Overexpression of OsDof12 affects plant architecture in rice (Oryza sativa L.)
The promoter of a H2O2- inducible, Arabidopsis glutathione S-transferase gene contains closely linked OBF-and OBP1-binding sites
Genome-wide characterization and expression analysis of the Dof gene family related to abiotic stress in watermelon
ATDof5.8 protein is the up-stream regulator of ANAC069 and is responsive to abiotic stress
甜瓜Dof家族全基因组鉴定与表达分析
DOI:10.16420/j.issn.0513-353x.2019-0115
[本文引用: 1]
基于甜瓜全基因组数据信息,利用生物信息学方法对甜瓜Dof家族基因进行了全面鉴定和系统发育分析,并通过Real-time PCR方法检测了CmDof家族成员对模拟干旱(PEG6000)、盐和低温胁迫的应答。结果表明:在甜瓜中存在34个Dof基因,除1、5和7号染色体外其余染色体均有分布,发生11次染色体片段重复事件和2次串联重复事件,外显子数在1 ~ 3之间,其蛋白质等电点在4.95 ~ 9.59之间;蛋白序列系统发育分析显示CmDof蛋白可分为9个进化亚群;组织表达模式分析发现该家族基因具有一定的时空表达特异性;多数CmDof基因对PEG6000、盐和低温胁迫有响应,其中CmDof3/CmDof5/CmDof10/ CmDof17、CmDof3/CmDof5/CmDof10和CmDof3/CmDof5/CmDof32分别响应PEG6000、盐和低温胁迫。
玉米Dof转录因子家族的全基因组鉴定与分析
DOI:10.3864/j.issn.0578-1752.2014.23.002
[本文引用: 1]
【目的】全基因组水平鉴定并解析玉米Dof(DNA binding with one finger)基因家族。【方法】基于玉米V3基因组数据鉴定玉米Dof基因家族,并从基因的结构、系统发育关系、染色体的位置分布、玉米不同组织和不同生理发育阶段基因的表达谱以及在充足氮(sufficient nitrogen,SN)和低氮(limiting nitrogen,LN)条件下V3期叶组织基因的差异表达5个方面分析玉米Dof基因家族。【结果】玉米参考基因组中存在46个Dof结构域基因,命名为ZmV3Dof1—ZmV3Dof46。通过系统发育关系和序列相似性将该基因家族分为8个亚类(Subgroup)S1—S8,每个亚类有3—8个Dof基因。通过分析该家族染色体分布,发现玉米10条染色体,除Chr.9外,均有Dof基因的分布,其中,在Chr.1上分布最为密集,有12个ZmV3Dofs;在Chr.5和Chr.3上分布次之,分别为8和7个ZmV3Dofs;在其他染色体上则分布较少,此外,Dofs在Chr.1的底部、Chr.5的顶部和底部分布密度相对较高。表达谱分析结果表明,Dof基因家族成员在不同组织及不同发育阶段均有差异表达,预示不同Dof基因功能的多样性以及在植物发育过程中扮演着不同角色,且同一亚类Dof基因表达模式存在一定程度的相似性。此外,玉米Dof基因家族对氮响应较为敏感,充足氮(SN)和低氮(LN)条件下,V3期叶组织基因的RNAseq表达结果显示35个Dof基因在2种氮处理下表达量存在差异,其中13个基因表达量差异较大。值得注意的是21个Dof基因在低氮处理下表达量上调,其中8个基因只在低氮条件下表达。【结论】基于最新版基因组数据鉴定玉米在自交系B73基因组中存在46个Dof基因,Dof基因家族成员参与玉米不同生理发育过程中,部分基因可能在氮代谢调控中发挥积极作用。
SbWRKY30 enhances the drought tolerance of plants and regulates a drought stress- responsive gene, SbRD19, in sorghum
Tomato SlPti5 plays a regulative role in the plant immune response against Botrytis cinerea through modulation of ROS system and hormone pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}