


烟碱,又称尼古丁,是一种存在于茄科植物的生物碱。在烟属植物中,不同品种资源烟碱含量差异较大。烤烟是目前世界上栽培面积最大的烟草类型,其烟碱含量中等,为1.5%~3.5%;白肋烟的烟碱含量高于烤烟,为2%~4%;香料烟烟碱含量较低,为1%~2%;雪茄烟烟碱含量为3%~5%;而黄花烟烟碱含量较高,在4%以上,最高可达8%。
经过近百年的研究,烟碱的合成途径及调控机理获得了较大的进展,烟碱合成途径已基本解析清楚。烟碱在烟草根部合成,通过木质部向地上部运输,储存在叶肉细胞中[1]。在烟草植株中,叶片烟碱含量最高,高于根部及茎部[2]。烟碱由吡啶环及吡咯烷环组成[3]。其中吡啶环部分来自于吡啶核苷酸循环途径中的烟酸,其前体是由天冬氨酸合成的喹啉酸,喹啉酸在喹啉酸磷酸核糖转移酶(quinoline phosphoribosyl transferase,QPT)催化下形成烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD),然后经由吡啶核苷酸循环途径生成烟酸[4]。烟碱的吡咯烷环来自于鸟氨酸,其在鸟氨酸脱羧酶的催化下形成腐胺,腐胺在腐胺N-甲基转移酶(putrescine-N-methyl transferase,PMT)作用下形成N-甲基腐胺,随后在N-甲基腐胺氧化酶(N-methylputrescine oxidase,MPO)催化下形成4-甲氨基丁醚,并自身环化形成N-甲基-∆1-吡咯啉阳离子,随后与烟酸(或其衍生物)发生缩合反应形成烟碱[5]。
烟碱的合成受多种因素调控,茉莉酸是目前研究较为深入的调控烟碱合成的植物激素,施用外源茉莉素或增加内源茉莉素水平都会诱导烟碱合成相关基因的快速表达[6-7]。茉莉酸在受伤诱导的烟碱合成中也扮演着重要角色[8-9]。茉莉酸途径参与烟碱合成调控的机理目前已研究的比较深入,茉莉酸可结合NtCOI蛋白,导致茉莉酸途径另一重要成员NtJAZ的降解,NtJAZ的降解释放了之前与之结合、丧失活性的转录因子蛋白NtHLH,活化的NtHLH进一步激活烟碱合成途径基因的表达[5]。通过调控茉莉酸途径成员,可以达到调控烟碱合成的效果。如沉默NtCOI1基因导致烟碱不能被茉莉酸及叶片受伤诱导。另外,缺失JAZ蛋白的JAS结构域使得JAZ蛋白不能被降解,导致茉莉酸诱导烟碱合成受到抑制[10]。MYC2是一种NtHLH类转录因子,参与了烟碱的合成调控。MYC2通过2种方式正向调控烟碱。一是直接作用于烟碱合成途径及转运基因的启动子区域上调基因的表达;二是间接作用于另一类烟碱合成调控因子ERF,进而调控烟碱的合成[11]。
甲基磺酸乙酯(EMS)是一种化学诱变剂,在作物育种中广泛应用,具有诱变效率高、诱变效果好、染色体畸变较少的优点。EMS诱变在植物育种上的应用,不仅可以打破传统的植物育种方式,获得的突变群体,还对培育植物新品种和丰富植物遗传资源具有重要意义。利用TILLING技术可以快速检测并精确获得目标基因发生突变的突变体材料,进入育种程序。本研究通过在我国主栽烤烟品种云烟87的EMS突变体库中筛选NtJAZ1基因突变体,获得烟碱含量升高且稳定遗传的突变体材料,为后续高烟碱烤烟新品种培育提供种质资源。
1 材料与方法
1.1 试验材料及试验地概况
云烟87的EMS突变体种植于云南省烟草农业科学研究院温室内。采用盆栽,塑料盆规格为33 cm×28 cm,昼/夜温度为25±1 ℃/18±1 ℃,每株施烟草专用肥(N:P:K=1:1.5:2.8)30 g,分5次施入。
1.2 EMS突变体种植及DNA提取
2019年温室种植云烟87的EMS突变体M2代1842份。在旺长期利用DNeasy Plant Mini Kit提取DNA。将突变体库DNA样品按照编号顺序排列,放置于96孔样品板内。分别取2 μL DNA样品进行浓度测定,将所有样品DNA浓度稀释至40 ng/μL,然后混合组成8倍样品混池用于TILLING检测。
1.3 NtJAZ1基因突变体植株的筛选
将NtJAZ1基因组序列(来自烟草行业内基因组数据库)提交在线软件PRIMER3,设计TILLING检测引物JAZ-TF(AAACTTGTAATCTTTCGAG CCAATTC)和JAZ-TR(ATCTGCTGGAAAATCAT CAAATACAA)。TILLING检测参照文献[12]并略作优化,具体步骤如下。
1.3.1 PCR扩增形成异源双链
PCR体系为10 μL:10×Buffer 1.0 μL,dNTP(2.5 mmol/L)0.8 μL,JAZ-F(10 μmol/L)0.16 μL,JAZ-R(10 μmol/L)0.16 μL,DNA 1.0 μL,灭菌ddH2O 6.88 μL。反应条件:95 ℃预变性3 min;95 ℃变性30 s,62 ℃退火30s(-1 ℃/循环),72 ℃延伸90 s,7个循环;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸90 s,40个循环;75 ℃延伸5 min;4 ℃保存。
1.3.2 Cel I酶切
用特异性切割错配碱基的核酸内切酶Cel I消化PCR形成的异源双链核酸分子。Cel I酶切体系体积为6 μL,包含0.2 μL Cel I(1个单位)、1.2 μL 10× Cel I Buffer、2 μL PCR产物和2.6 μL去离子水。采用AdvanCE FS96(Advanced Analytical Technologies,美国)进行毛细管电泳,筛选获得突变体单株。
1.3.3 突变单株测序确证
为了确证突变体位点,设计突变位点检测引物JAZdF(ACAGGAAACCA AACAACTACAACT)和JAZdR(CCCAGTACACT AAGCTTCTGCTAT)。使用Q5 DNA聚合酶(NEB)进行PCR反应,反应程序如下:98 ℃预变性30 s;98 ℃变性7 s,60 ℃退火30 s,72 ℃延伸30 s,30个循环;72 ℃延伸5 min,4 ℃保存。PCR产物经纯化后用送赛默飞世尔科技(中国)有限公司(Invitrogen)测序,测序引物为JAZdF。对突变个体PCR产物进行克隆测序,鉴定突变类型和位置。
1.4 M4代突变体材料及后代烟碱含量检测
2020年种植突变体M3代株系,通过PCR扩增、产物测序鉴定获得纯合突变体株系。温室播种筛选获得的11个突变体纯合株系M4代,待植株生长至现蕾期,取全株烟叶,于60 ℃烘干并参照YC/T 383-2010[13]测定烟碱含量。烟碱含量显著提高的突变体自交收种。
2021年温室种植J11后代2个株系(J11 I-1和J11 Ⅳ-1)和云烟87各20株,中心花开放期打顶,打顶前取10株整株烟叶,打顶后2周取10株整株烟叶,于60 ℃烘干并参照YC/T 383-2010[13]测定烟叶烟碱含量。另外打顶前取J11 I-1和云烟87根部样品对NtPMT1a和NtQPT基因的表达进行分析,进行3次生物学重复。NtPMT1a基因qPCR引物为PMT1a-F(AAATGGCACTTCTGAACACCTC)和PMT1a-R(CCCATTCTGGTGGCCGTTCC)。NtQPT基因qPCR引物为QPT-F(TACAAGAGTGG AGTCATTAGAG)和QPT-R(GCAAGTGCAAT TCTGCTATG)。
2 结果与分析
2.1 突变体材料的获得
图1
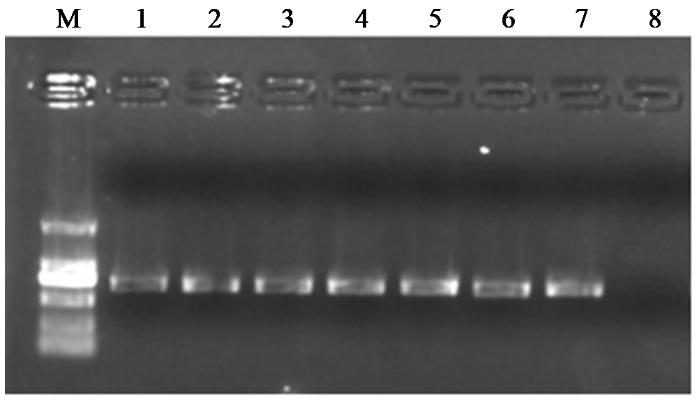
图1
突变位点检测引物PCR扩增结果
Fig.1
PCR amplification results of mutation site detection primers
图2
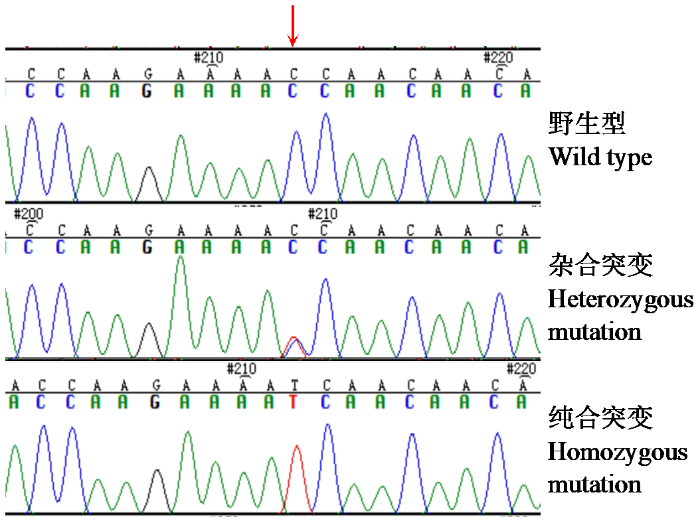
图2
J11株系3个基因型
箭头所示为突变碱基位置。
Fig.2
Three genotypes of J11 lines
Arrows indicate mutant base locations.
表1 筛选获得的NtJAZ1基因突变体信息
Table 1
| 突变体编号 Number of mutants | 核苷酸突变 Nucleotide mutation | 氨基酸突变体 Amino acid mutation | 突变位点所在结构域 The domain of the mutation site |
|---|---|---|---|
| J2 | C305T | T102I | ZIM |
| J3 | C346T | P116S | ZIM |
| J4 | G584A | R195K | Jas |
| J5 | C650T | T217I | C端 |
| J6 | G652A | E218K | C端 |
| J8 | G500A | R167Q | ZIM和Jas间隔区 |
| J9 | C708T | A203V | Jas |
| J10 | C616T | Q206* | Jas |
| J11 | C392T | T131I | ZIM和Jas间隔区 |
| J14 | C473T | T158I | ZIM和Jas间隔区 |
| J15 | G372A | M124I | ZIM |
| J16 | 内含子 | 内含子 |
“*”代表突变位点位于内含子。
“*”indicates that the mutation site is located in intron.
图3
2.2 M4代纯合突变体材料烟碱含量
温室盆栽种植筛选获得的M4代纯合突变株系,现蕾期取整株叶片,杀青烘干后检测烟碱含量。结果(图4)表明,J8和J11烟碱含量显著高于野生型材料,除了J6明显低于对照外,其他突变体株系烟碱含量与对照差异不明显。J8和J11突变位点均位于ZIM和Jas结构域间隔区,而位于ZIM和Jas结构域内的突变烟碱含量反而没有明显提高。
图4
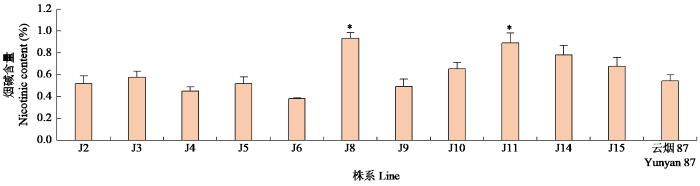
图4
不同位点纯合突变体株系烟叶烟碱含量
“*”表示在P < 0.05水平差异显著。下同。
Fig.4
Nicotine contents in tobacco leaves of homozygous mutant strains with different loci
“*”indicates significant difference at P < 0.05 level. The same below.
J11突变体的突变类型为NtJAZ1基因CDS区第392位的核苷酸由C变为T,导致NtJAZ1蛋白氨基酸残基第131位的苏氨酸(T)突变为异亮氨酸(I)。T属于极性氨基酸,I属于非极性氨基酸,T变为I可能对蛋白质的三维结构造成影响。
2.3 J11后代材料烟碱含量及NtPMT1a和NtQPT基因表达水平分析
从J11收获2个株系J11 I-1和J11 IV-1(M5代)。温室种植J11 I-1和J11 IV-1,研究其高烟碱表型是否可以稳定遗传。由图5~6可知,打顶前J11 I-1和J11 IV-1的烟碱含量分别为0.50%和0.49%,极显著高于对照云烟87(0.31%),较对照分别提高约60%和55%。打顶后2个株系的烟碱含量(0.89%、0.95%)也极显著高于对照云烟87(0.60%),较对照分别提高约48%和58%。PMT和QPT是烟碱合成途径中的2个限速酶,对J11I-1打顶前根部NtPMT1a和QPT基因的表达水平进行了检测,结果(图7)显示,2个基因在突变体中的表达水平极显著提高。
图5
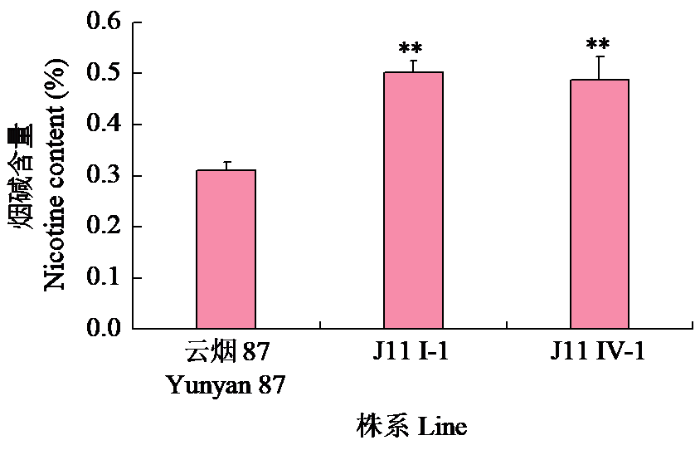
图5
打顶前温室J11 M4代2个株系烟碱含量
“**”表示在P < 0.01水平差异显著。下同。
Fig.5
Nicotine content in two lines of J11 M4 generation in greenhouse before topping
“**”indicates significant difference at P < 0.01 level. The same below.
图6
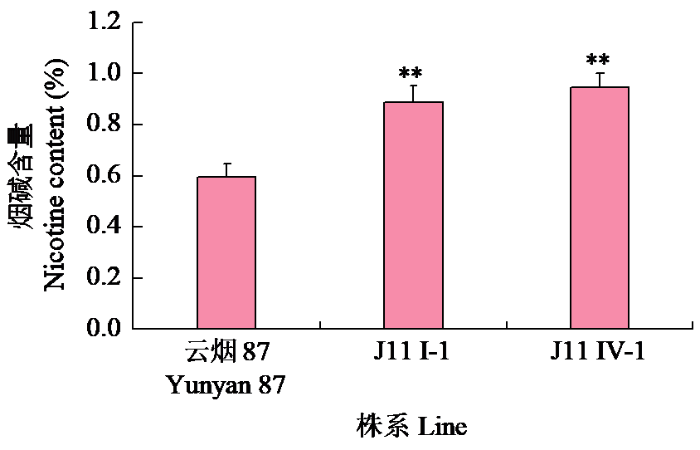
图6
打顶后温室J11 M4代2个株系烟碱含量
Fig.6
Nicotine content in two lines of J11 M4 generation in greenhouse after topping
图7
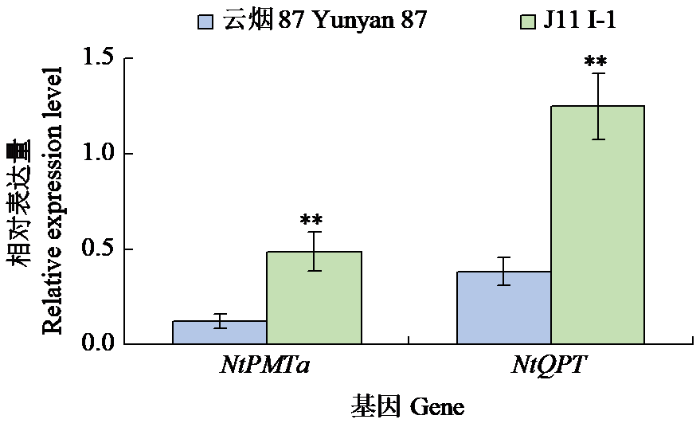
图7
打顶前J11 I-1 NtPMT1a和NtQPT基因的表达水平
Fig.7
Expression level of NtPMT1a and NtQPT genes in J11 I-1 line before topping
3 讨论
烟碱是烟草中重要的化学物质。目前,通过工业降焦产生的低焦油卷烟会引起烟气烟碱含量降低,不能很好地满足市场需求。农业生产上可通过大水大肥措施获得高烟碱的烟叶,但水肥过量会导致烟叶难烘烤,品质下降。通过转基因手段也可获得烟叶烟碱含量提高的烟草品系,如过表达NtMYC2a基因可以使烟叶烟碱含量提高70%左右[14],但转基因材料目前还不被市场接受。为了满足卷烟工业企业对高烟碱烟叶原料的需求,本研究以EMS诱变为育种手段,以烟碱负调控因子基因NtJAZ1为目标,对主栽品种云烟87的EMS突变体库进行筛选,辅以分子标记选择,获得了烟碱含量提高约50%的烤烟新品系,为培育高烟碱新品种提供了资源。
JAZ1蛋白包含ZIM和Jas 2个保守结构域。ZIM结构域在JAZ蛋白同源及异源二聚体的形成过程中发挥作用[15],而Jas结构域与多个转录因子蛋白的结合,如MYC2、DELLA蛋白等,从而参与调控多种激素的信号传导途径[16]。在茉莉酸存在的情况下,NtJAZ1蛋白降解后释放与之结合的MYC2[17],具有活性的MYC2可促进烟碱的合成[11]。从以上的调控机理推测,Jas结构域中的突变有可能影响NtJAZ1与MYC2a的互作,从而引起烟碱合成水平发生改变。本研究获得3个位于Jas结构域中的突变体株系(J4、J9和J10),其中J4由精氨酸转变为赖氨酸,J9由丙氨酸转变为缬氨酸。以上2种转换都是同类氨基酸之间的转换,J4和J9突变体烟碱含量没有显著变化,推测这些突变对NtJAZ1蛋白功能影响不大;J10为谷氨酰胺突变为终止密码子,产生终止突变,但并没有影响烟碱的合成,暗示NtJAZ1在调控烟草烟碱合成方面可能存在复杂的机制[18-19]。
本研究从云烟87突变体库中筛选获得11个突变体材料,这些突变体突变位点位于NtJAZ1蛋白的不同区域。烟碱含量显著提高的J8及J11的突变位点均位于ZIM及Jas结构域间隔区,其他区域中产生的突变并不能引起烟碱含量的提高。推测ZIM及Jas结构域间隔区可能对NtJAZ1三维结构的形成具有关键作用,当然,该推断还需要进一步试验验证。以上试验结果也表明,不同JAZ1蛋白区域在该蛋白行使调控烟碱合成的功能中发挥着不同的作用。
栽培烟草是异源四倍体,基因组中含有2套基因组,包含多个同源家族成员。研究[17]发现,烟草基因组中含有15个JAZ家族成员,部分成员之间在功能上存在冗余的可能性。本研究通过突变NtJAZ家族一个成员获得烟碱含量提高约50%的烤烟新品系,如果期望获得烟碱含量更高的烟草材料,推测需同时突变多个JAZ家族基因来实现。
4 结论
在烟草主栽品种云烟87 EMS突变体中筛选烟碱合成负调控基因NtJAZ1的突变体,获得10个氨基酸发生改变的突变体,1个无义突变体。在纯合突变体后代筛选获得2个突变体株系,其烟碱含量显著升高。对其中1个株系后代烟碱含量分析表明其高烟碱表型可以稳定遗传,且该株系中烟碱合成关键酶基因NtPMT1a和NtQPT表达水平显著升高。该研究获得的高烟碱材料可为培育高烟碱烤烟新品种提供种质资源。
参考文献
Nicotine synthesis in excised roots
Accumulation of nicotine in reciprocal grafts of tomato and tobacco
Molecular regulation of nicotine biosynthesis
Regulation in tobacco callus of enzyme activities of the nicotine pathway II. The pyridine- nucleotide cycle
DOI:10.1007/BF00392369
PMID:24232153
[本文引用: 1]

In tobacco callus, the induction of nicotine synthesis, which stimulates enzyme activities of the ornithine-methylpyrroline route (see the preceding paper), also leads to marked changes in the enzyme activities of the pyridine-nucleotide cycle. This cycle provides the metabolite (probably nicotinic acid) for condensation with methylpyrroline to produce nicotine. The activities of eight enzymes of the pyridine-nucleotide cycle and of quinolinic-acid phosphoribosyltransferase, the anaplerotic enzyme, were determined by high-performance liquid chromatography assays. The distinct changes of their activities upon induction of nicotine synthesis lead to the following conclusions: i) nicotinic acid is the relevant metabolite which is provided by the pyridine-nucleotide cycle and consumed for nicotine synthesis. ii) The enhancement of the nicotinic-acid pool arises in two ways, by synthesis of NAD and degradation via nicotinamide mononucleotide and by a direct route from nicotinic-acid mononucleotide (NaMN) which is degraded by a glycohydrolase with a rather high K m value. Such a K m value prevents the complete depletion of the NaMN pool.
Molecular genetics of alkaloid biosynthesis in Nicotiana tabacum
DOI:10.1016/j.phytochem.2013.06.002
PMID:23953973
[本文引用: 2]
Alkaloids represent an extensive group of nitrogen-containing secondary metabolites that are widely distributed throughout the plant kingdom. The pyridine alkaloids of tobacco (Nicotiana tabacum L.) have been the subject of particularly intensive investigation, driven largely due to the widespread use of tobacco products by society and the role that nicotine (16) (see Fig. 1) plays as the primary compound responsible for making the consumption of these products both pleasurable and addictive. In a typical commercial tobacco plant, nicotine (16) comprises about 90% of the total alkaloid pool, with the alkaloids nornicotine (17) (a demethylated derivative of nicotine), anatabine (15) and anabasine (5) making up most of the remainder. Advances in molecular biology have led to the characterization of the majority of the genes encoding the enzymes directly responsible the biosynthesis of nicotine (16) and nornicotine (17), while notable gaps remain within the anatabine (15) and anabasine (5) biosynthetic pathways. Several of the genes involved in the transcriptional regulation and transport of nicotine (16) have also been elucidated. Investigations of the molecular genetics of tobacco alkaloids have not only provided plant biologists with insights into the mechanisms underlying the synthesis and accumulation of this important class of plant alkaloids, they have also yielded tools and strategies for modifying the tobacco alkaloid composition in a manner that can result in changing the levels of nicotine (16) within the leaf, or reducing the levels of a potent carcinogenic tobacco-specific nitrosamine (TSNA). This review summarizes recent advances in our understanding of the molecular genetics of alkaloid biosynthesis in tobacco, and discusses the potential for applying information accrued from these studies toward efforts designed to help mitigate some of the negative health consequences associated with the use of tobacco products. Copyright © 2013 Elsevier Ltd. All rights reserved.
Jasmonate-induced responses are costly but benefit plants under attack in native populations
Proceedings of the National Academy of Sciences of the United States of America,
Methyl jasmonate induced expression of the tobacco putrescine N-methyltransferase genes requires both G-box and GCC-motif elements
Wound-induced changes in root and shoot jasmonic acid pools correlate with induced nicotine synthesis in Nicotiana sylvestris Spegazzini and Comes
DOI:10.1007/BF02066250
PMID:24242736
[本文引用: 1]
Leaf damage by herbivores inNicotiana sylvestris Spegazzini and Comes (Solanaceae) produces a damage signal that dramatically increasesde novo nicotine synthesis in the roots. The increased synthesis leads to increases in whole-plant nicotine pools, which in turn make plants more resistant to further herbivore attack. Because signal production and the response to the signal occur in widely separated tissues, the speed with which different damage signals exit a damaged leaf can be studied. We propose that electrical damage signals should exit a leaf faster (less than 60 min) than chemical damage signals. Excision of a leaf induces a smaller increase in nicotine production than does puncture damage, so we examined our proposition by excising previously punctured leaves at 1, 60, and 960 min after leaf puncture and quantifying the induced whole-plant nicotine pools six days later when the induced nicotine production had reached a maximum. Significant induced nicotine production occurred only if punctured leaves were excised more than 1 hr after puncture, which is consistent with the characteristics of a slow-moving chemical signal rather than a fast-moving electrical signal. We explore the nature of the chemical signal and demonstrate that additions of 90µg or more of methyl jasmonate (MJ) in an aqueous solution to the roots of hydroponically grown plants inducede novo nicotine synthesis from(15)NO3 in a manner similar to that induced by leaf damage. We examine the hypothesis that jasmonic acid (JA) functions in the transfer of the damage signal from shoot to root. Using GC-MS techniques to quantify whole-plant JA pools, we demonstrate that leaf damage rapidly (<0.5 hr) increases shoot JA pools and, more slowly (<2 hr), root JA pools. JA levels subsequently decay to levels found in undamaged plants within 24 hr and 10 hr for shoots and roots, respectively. The addition of sufficient quantities (186µg) of MJ in a lanolin paste to leaves from hydroponically grown plants significantly increased endogenous root JA pools and increasedde novo nicotine synthesis in these plants. However, the addition of 93µg or less of MJ did not significantly increase endogenous root JA pools and did not significantly affectde novo nicotine synthesis. We propose that wounding increases shoot JA pools, which either directly through transport or indirectly through a systemin-like signal increase root JA pools, which, in turn, stimulate root nicotine synthesis and increase whole-plant nicotine pools.
Jasmonate induction of putrescine N-methyltransferase genes in roots of Nicotiana sylvestris
DOI:10.1093/pcp/pcd001
PMID:10965939
[本文引用: 1]
Nicotine alkaloids are synthesized in the root of Nicotiana species, and their synthesis increases after insect attack, wounding and jasmonate treatment of the leaf. Putrescine N-methyltransferase (PMT) catalyzes the first committed step in nicotine biosynthesis. The expression patterns of the three Nicotiana sylvestris PMT genes (NsPMT1, NsPMT2, and NsPMT3) are reported in this study. Transcripts of the NsPMT genes were detected only in the root, and were up-regulated by methyl jasmonate treatment. When the 5'-flanking regions of NsPMT1, NsPMT2, and NsPMT3 were fused independently to beta-glucuronidase reporter gene and introduced into N. sylvestris by Agrobacterium-mediated transformation, all introduced transgenes were expressed in the cortex, endodermis, and xylem in the root, as well as upregulated by methyl jasmonate treatment. These qualitatively similar patterns of expression for the NsPMT genes are achieved with only 0.25 kb of their conserved 5'-flanking regions, which contained no known jasmonate-responsive elements.
Jasmonate-induced nicotine formation in tobacco is mediated by tobacco COI1 and JAZ genes
Tobacco MYC2 regulates jasmonate- inducible nicotine biosynthesis genes directly and by way of the NIC2-locus ERF genes
A simple CELI endonuclease- based protocol for genotyping both SNPs and InDels
Genetic factors for enhancement of nicotine levels in cultivated tobacco
DOI:10.1038/srep17360
PMID:26626731
[本文引用: 1]
Nicotine has practical applications relating to smoking cessation devices and alternative nicotine products. Genetic manipulation for increasing nicotine content in cultivated tobacco (Nicotiana tabacum L.) may be of value for industrial purposes, including the possibility of enhancing the efficiency of nicotine extraction. Biotechnological approaches have been evaluated in connection with this objective, but field-based results are few. Here, we report characterization of two genes encoding basic-helix-loop-helix (bHLH) transcription factors (TFs), NtMYC2a and NtMYC2b from tobacco. Overexpression of NtMYC2a increased leaf nicotine levels in T-1 transgenic lines approximately 2.3-fold in greenhouse-grown plants of tobacco cultivar 'NC 95'. Subsequent field testing of T-2 and T-3 generations of transgenic NtMYC2a overexpression lines showed nicotine concentrations were 76% and 58% higher than control lines, respectively. These results demonstrated that the increased nicotine trait was stably inherited to the T-2 and T-3 generations, indicating the important role that NtMYC2a plays in regulating nicotine accumulation in N. tabacum and the great potential of NtMYC2a overexpression in tobacco plants for industrial nicotine production. Collected data in this study also indicated a negative feedback inhibition of nicotine biosynthesis. Further enhancement of nicotine accumulation in tobacco leaf may require modification of the processes of nicotine transport and deposition.
The ZIM domain mediates homo- and heteromeric interactions between Arabidopsis JAZ proteins
Transcriptome profiling identified multiple jasmonate ZIM-domain proteins involved in the regulation of alkaloid biosynthesis in tobacco BY-2 cells
The JAZ family of repressors is the missing link in jasmonate signalling
NtAIDP1, a novel NtJAZ interacting protein, binds to an AT-rich region to activate the transcription of jasmonate-inducible genes in tobacco
JASMONATE-ZIM DOMAIN proteins engage Polycomb chromatin modifiers to modulate Jasmonate signaling in Arabidopsis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}