


盐碱胁迫作为主要的非生物胁迫因素之一[7],会使植物表型发生变化,并对其生长发育造成不同程度的影响[8-9]。这些变化与相关代谢物表达量的变化密切相关[10-
1 材料与方法
1.1 试验材料与试验设计
以Xin-17和草莜1号为试验材料。挑选籽粒完整、大小均匀的种子,用0.1% HgCl2消毒8 min,无菌蒸馏水清洗3次,自然晾干。将燕麦种子种植于盛满蛭石的方形盆中(20 cm×15 cm×10 cm),播种深度约5 cm,每盆播种同品种50粒。将NaCl、Na2SO4、Na2CO3和NaHCO3按等摩尔比混合,配制0(pH 6.88,CK)、20(pH 7.48,T1)、30(pH 7.85,T2)、40(pH 8.07,T3)、50(pH 8.46,T4)和60 mmol/L(pH 8.88,T5)的复合盐碱溶液,分别取不同浓度盐碱溶液浇灌处理。每个处理设置6个重复,于温室中培养,自然光照,湿度约60%。出苗完成后,定期浇灌蒸馏水或1/4 Hoagland溶液。
1.2 表型及生理指标测定
播种后第6、7、8及13天测量各处理组的株高(每组取约20株)。播种后第8天统计各处理组的出苗率。第13天测量各处理组幼苗的干重和鲜重,并计算含水量。称取0.5 g叶片,剪碎并浸泡于95%异丙醇溶液中,4 ℃过夜,测量对应的吸光值,计算光合色素叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)的含量[26]。
收集3个盐碱浓度0(CK)、30(T2)和60 mmol/L(T5)处理的叶片组织,立即用液氮冷冻,保存于-80 ℃冰箱。每个处理分别取1.0 g组织样本于液氮中快速研磨成粉末,用试剂盒TE0741(北京雷根生物技术有限公司)测定过氧化氢酶(CAT)活性,用试剂盒TE0727(北京雷根生物技术有限公司)测定超氧化物歧化酶(SOD)活性。
1.3 组织代谢物提取
收集CK、T2和T5处理的叶片组织,每个处理设置6个重复,取样后立即用液氮冷冻,保存于-80 ℃冰箱。每个处理分别取100 mg组织样本于液氮中研磨,然后置于EP管中,加入500 μL含0.1%甲酸的80%甲醇水溶液,涡旋震荡,冰浴静置5 min,15 000转/min、4 ℃离心10 min,取一定量的上清液加质谱级水稀释,并置于离心管中15 000转/min、4 ℃离心10 min,收集上清,继续下一步试验。从每个试验样本中取等体积样本混匀作为QC(quantity control)样本。
1.4 LC-MS/MS分析及代谢物鉴定
对处理后各样品进行LC-MS/MS分析。色谱柱为Hypersil Gold column(C18),柱温设定为40 ℃,进样流速为0.2 mL/min。正模式的流动相A为0.1%甲酸,流动相B为甲醇;负模式的流动相A为5 mmol/L醋酸铵,pH 9.0;流动相B为甲醇。质谱扫描范围选择m/z 70-1050。一级质谱分辨率为70 000(200 m/z),AGC(automatic gain control)为1×106,maximum IT为50 ms,动态排除(dynamic exclusion)为60.0 s。
将质谱检测得到的原始文件(.raw)导入Compound Discoverer 3.1(CD)软件中,进行保留时间和质荷比等参数的筛选,将不同样品根据保留时间偏差0.2 min和质量偏差0.0005%进行峰对齐。根据设置的质量偏差0.0005%、信号强度偏差30%、信噪比3、最小信号强度100 000以及加和离子等信息进行峰提取。然后通过高分辨XIC图中的质荷比确定化合物的精确分子量,根据质量数偏差和加合物离子信息进行分子式预测。同时,对峰面积进行定量,整合目标离子,通过对分子、离子峰和碎片离子进行分子式的预测并与mzCloud、mzVault和Mass List数据库进行比对,对分子特征峰进行匹配,通过与mzCloud数据库中每个化合物的碎片离子和碰撞能等信息进行匹配,对体系中的代谢物进行鉴定。然后选取QC样本中变异系数(coefficient of variance,CV)值小于30%的化合物作为最终鉴定结果。用Blank样本去除背景离子,并对定量结果进行归一化,最后完成数据的鉴定和定量。
1.5 差异代谢物分析
对代谢物数据进行多元统计分析,将所有试验样本和QC样本中提取得到的峰,经预处理后进行PCA分析。采用PLS-DA模型筛选差异代谢物,其中差异倍数(fold change,FC)定义为比较组中各代谢物定量均值的比值,并计算其以2为底的对数(log2FC)。差异代谢物筛选标准为PLS-DA模型第1主成分的变量投影重要度(variable importance in the projection,VIP)值大于1.0,t-检验的P值小于0.05,且FC值大于1.5或小于0.667,即|log2FC|>log2(1.5)。利用KEGG(kyoto encyclopedia of genes and genomes)数据库进行功能注释及数据分析,确定代谢物参与的生化代谢途径,通过富集分析在差异代谢物中确定主要的代谢途径,显著性标准设置为P<0.05,q<0.05。
1.6 qRT-PCR检测
收集CK、T2和T5处理的叶片组织,迅速在液氮中研磨成粉末,使用RNA提取试剂盒[天根生化科技(北京)有限公司,DP432]提取总RNA。使用Nano Drop(Thermo Fisher)测定RNA浓度,取3 μL提取的总RNA样,用1%琼脂糖凝胶电泳检测总RNA的完整性和纯度。每个样品取1 μg总RNA,用试剂盒[天根生化科技(北京)有限公司]去除DNA,利用cDNA反转录试剂盒[天根生化科技(北京)有限公司]合成cDNA,置于-20 °C保存备用。
根据燕麦基因组数据,选取了6个代谢物合成关键基因,设计引物后分析基因表达量,以β-actin作为内参(表1)。扩增反应体系:Taq PCR Master Mix 10 μL,10 μmol/L正向引物0.5 μL,10 μmol/L反向引物0.5 μL,cDNA模板1 μL,8 μL ddH2O,总体系20 μL。扩增程序:95 °C预变性3 min;95 °C变性10 s,60 °C退火30 s,72 °C延伸30 s,40个循环。1%琼脂糖凝胶电泳检测。每个样品3次技术重复。使用荧光定量试剂盒Super Real PreMix Plus[天根生化科技(北京)有限公司]进行试验,利用ABI QuantStudio 7 Flex实时荧光定量PCR系统检测。对基因的扩增曲线及熔解曲线进行分析,使用2-∆∆ct法分析数据。
表1 qRT-PCR检测基因及对应引物
Table 1
| 编号Code | 基因ID Gene ID | 基因功能注释Gene function annotation | 引物序列(5′-3′)Primer sequence (5′-3′) |
|---|---|---|---|
| 1 | Cluster-38721.80008 | YUCCA,吲哚-3-丙酮酸单加氧酶 | GCTGCCTGCTGATGTCTTGGG |
| GTTCGGCGACAACGGGATGG | |||
| 2 | Cluster-38721.50882 | 苯丙氨酸解氨酶 | TGAGGAGGAGCTTCGACAGACAC |
| GGTACAGAGGGTATGAACGGCAATC | |||
| 3 | Cluster-38721.77689 | 硝酸根高效转运蛋白2.2 | CGACCCAGCCAGAAGCAGAATG |
| GACCTTGGTGCCCGCTACTTTG | |||
| 4 | Cluster-38721.84959 | 磷转运蛋白2 | GCCCCTGCCCATATGCTAAA |
| TTATGGCCACGAGCATGACA | |||
| 5 | Cluster-38721.55805 | 淀粉合酶 | CTTGGTCGTCCCGTGAAAGA |
| ATGCCTCAGTGGAGCAGAAC | |||
| 6 | Cluster-38721.47252 | 蔗糖合成酶 | GCACCGTGAGTATCAGCGATGTAG |
| ACCTGTTTGGGCAGTTCCGTTG | |||
| 7 | - | β-肌动蛋白(内参基因) | CCAGCGAGGTCAAGACGAA |
| CCCAAGGCTAACAGGGAGAA |
1.7 数据处理
表型、生理指标以及qRT-PCR数据均为均值±SD值。使用SigmaPlot 14.0软件进行差异显著性分析(P<0.01),使用Excel 2010软件作图。
2 结果与分析
2.1 复合盐碱胁迫对燕麦幼苗生长的影响
图1
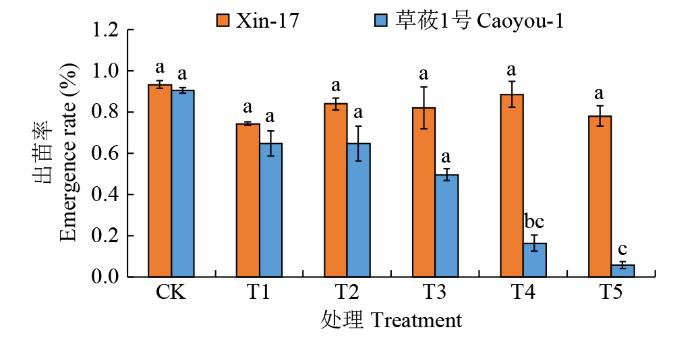
图1
复合盐碱胁迫下燕麦的出苗率
不同小写字母表示P < 0.05水平差异显著,下同。
Fig.1
Emergence rate of oats under compound saline-alkali stress
Different lowercase letters indicate significant differences at the P < 0.05 level, the same below.
图2

图2
复合盐碱胁迫下燕麦苗期的生长情况
Fig.2
Growth of oat at seedling stage under compound saline-alkali stress
图3
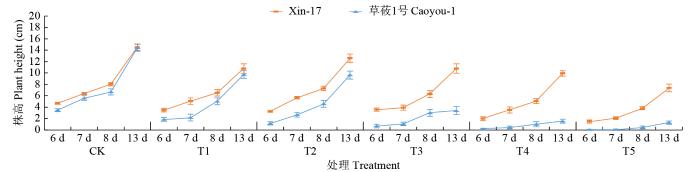
图3
复合盐碱胁迫下第6、7、8和13天的燕麦苗期株高变化
Fig.3
Changes of oat seedling plant height on the 6th, 7th, 8th, and 13th days under compound saline-alkali stress
2.2 复合盐碱胁迫对燕麦幼苗生理的影响
复合盐碱胁迫对燕麦幼苗的地上部干重和含水量有明显影响,但2个品种同样表现出明显差异(图4)。具体表现为,随着胁迫浓度的增加,草莜1号的含水量在40 mmol/L(T3)后逐渐减少,而相同胁迫浓度下的地上部干重有细微增加趋势;Xin-17的含水量随着胁迫浓度增加表现为先增加且超越草莜1号,后减少至接近草莜1号的水平;其地上部干重则为先降低后增加的趋势。2个品种的地上部干重和含水量变化趋势显示出抗性品种Xin-17与敏感品种草莜1号应对盐碱胁迫不同的适应机制,抗性品种维持较高的水分含量将有利于其在盐碱胁迫环境下的代谢,这与对应植株的表型变化一致。
图4
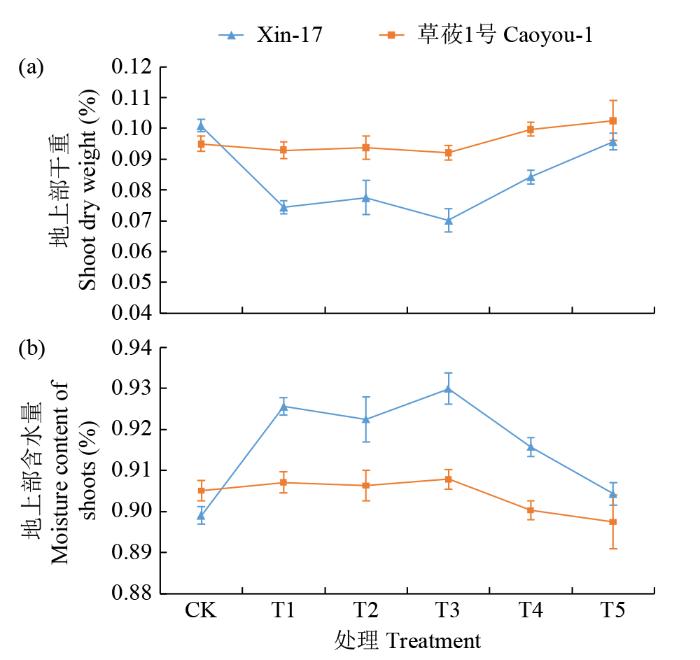
图4
复合盐碱胁迫下燕麦苗期地上部干重及含水量的变化
Fig.4
Shoot dry weight and moisture content changes of oat at seedling stage under compound saline-alkali stress
进一步分析盐碱胁迫对燕麦苗期光合色素含量的影响发现,复合盐碱胁迫影响了2个品种的3种光合色素(Chla、Chlb和Car)的含量,同时品种间也表现出明显的差异(图5)。随着盐碱浓度的增加,草莜1号中的3种光合色素含量均呈现出小幅度的下降趋势,而Xin-17中3种光合色素的含量随着盐碱浓度的增加,均表现为明显的下降趋势。2个品种间光合色素含量有明显差异,CK处理下Xin-17光合色素含量明显高于草莜1号,前者同等质量组织中光合色素含量是后者的3倍多,虽然随着胁迫浓度增加Xin-17中光合色素含量明显下降,但T5处理时仍然高于草莜1号。高浓度的光合色素有利于抗性品种维持稳定高效的光合作用,也有利于其抵御盐碱胁迫。
图5
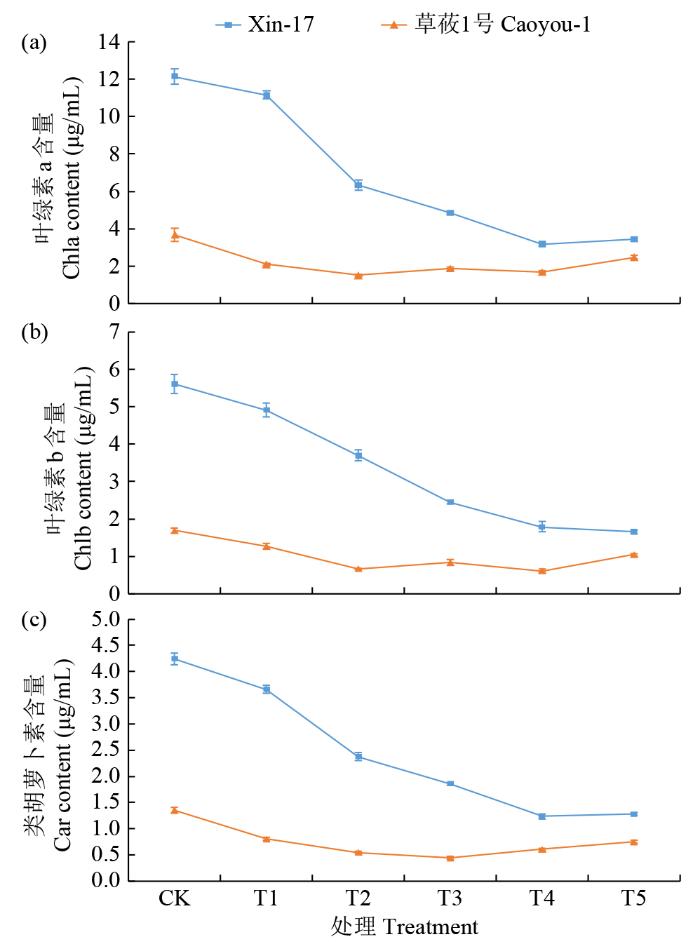
图5
复合盐碱胁迫下燕麦苗期光合色素含量的变化
Fig.5
Photosynthetic pigment content changes of oat at seedling stage under compound saline-alkali stress
对盐碱胁迫下2个品种中CAT和SOD活性的变化进行研究(图6)。在T2处理下,Xin-17中未检测到明显变化,而草莜1号中CAT活性显著增加;当浓度升高至60 mmol/L(T5),2个品种中的CAT和SOD活性均显著增加,Xin-17增加幅度更大。
图6
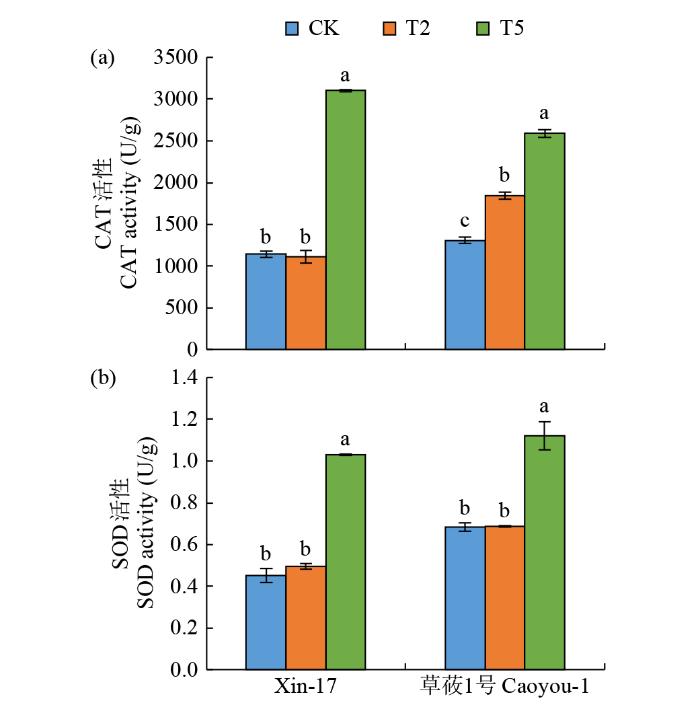
图6
复合盐碱胁迫下燕麦苗期CAT和SOD活性的变化
Fig.6
CAT and SOD activity changes of oat at seedling stage under compound saline-alkali stress
2.3 复合盐碱胁迫对燕麦苗期代谢组的影响
为了更详尽地解析复合盐碱胁迫对燕麦苗期的影响,对Xin-17和草莜1号的3个浓度盐碱处理0 mmol/L(CK)、30 mmol/L(T2)和60 mmol/L(T5)进行了代谢组学研究。经过LC-MS/MS检测、鉴定并注释,共得到973个代谢物,C1T5vs C1CK、C1T2 vs C1CK、X17T5 vs X17CK和X17T2 vs X17CK共4个组分别得到212、176、198和182个差异代谢物。对所有试验样本和QC样本进行PCA分析,椭圆为95%的置信区间(图7)。PCA分析发现,盐碱处理组(C1T2,C1T5,X17T2,X17T5)与对照组(C1CK,X17CK)明显分离,盐碱胁迫效果明显。
图7
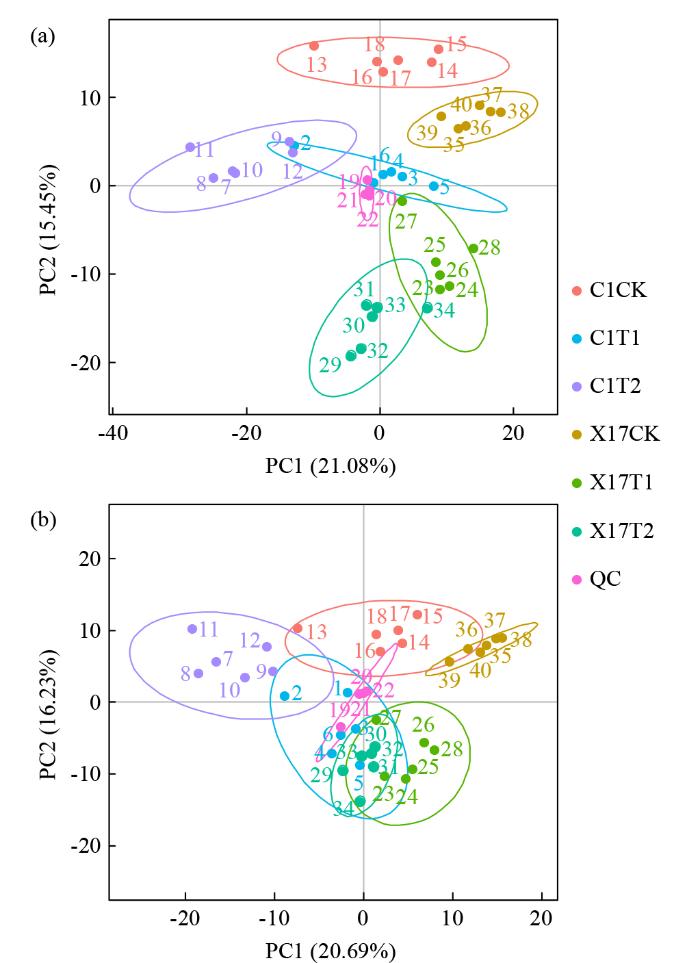
图7
复合盐碱胁迫下燕麦苗期PCA分析图
(a) 阳离子;(b) 阴离子。
Fig.7
PCA plot of oat at seedling stage under compound saline-alkali stress
(a) Cation; (b) Anion.
KEGG富集分析结果(图8)发现,盐碱胁迫浓度从30 mmol/L(T2)增加到60 mmol/L(T5),草莜1号中富集得到的Top20代谢通路中的差异代谢物总数从87个减少到34个;而Xin-17富集得到的Top20代谢通路的差异代谢物总数从48个增加到67个。2个品种各自的Top20代谢通路多数相同,如嘌呤及嘧啶代谢类、磷酸盐和亚磷酸盐代谢和硫代谢等;同时,2个品种富集得到的Top20代谢通路也存在明显不同,如Xin-17中富集得到ABC转运蛋白、卟啉和叶绿素代谢、脂肪酸延长、丙酸代谢和果糖与甘露糖代谢等,而草莜1号富集得到黄酮和黄酮醇生物合成、戊糖磷酸途径及戊糖转化、烟酸和烟酰胺代谢等。上述富集得到Top20代谢通路的特征性差异,可以明显体现出抗性品种与敏感品种的代谢差异。
图8
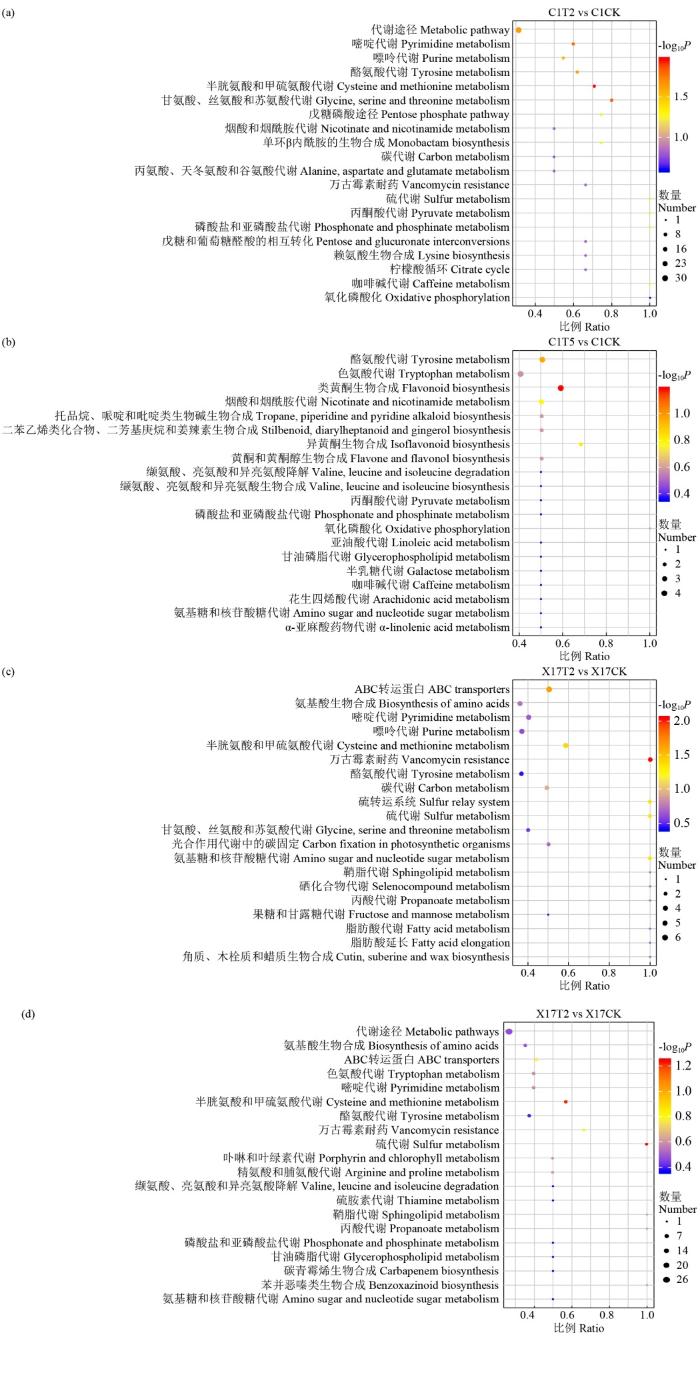
图8
复合盐碱胁迫下燕麦苗期主要代谢通路的KEGG富集分析(Top20代谢途径)
Fig.8
KEGG metabolites enrichment analysis of oat at seedling stage under compound saline-alkali stress (Top20 metabolic pathways)
2.4 复合盐碱胁迫对苗期燕麦色氨酸及次生代谢通路的影响
图9
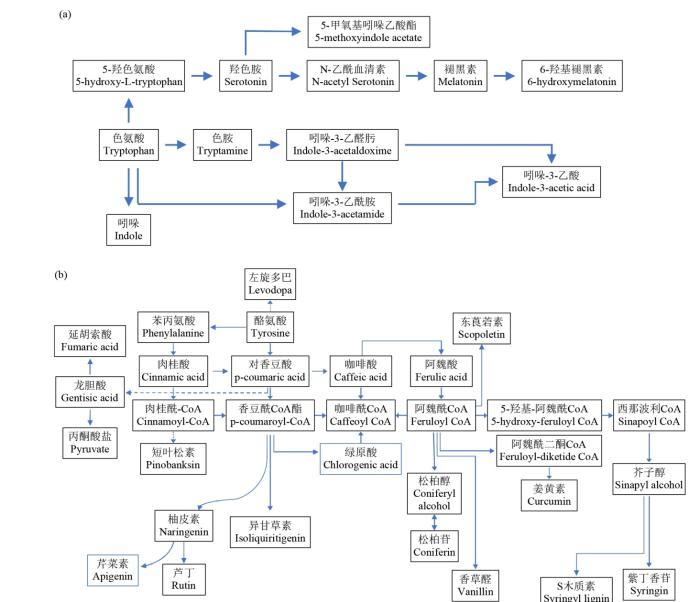
图9
复合盐碱胁迫下Xin-17和草莜1号的色氨酸代谢通路(a)和次生代谢通路(b)
Fig.9
Tryptophan metabolic pathway (a) and secondary metabolic pathway (b) of Xin-17 and Caoyou-1 under compound saline-alkali stress
表2 复合盐碱胁迫下色氨酸代谢通路与次生代谢通路中的主要代谢物
Table 2
| 代谢物ID Metabolite ID | 代谢物 Metabolite | X17CK vs C1CK | C1T2 vs C1CK | C1T5 vs C1CK | X17T2 vs X17CK | X17T5 vs X17CK | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| log2FC | P | log2FC | P | log2FC | P | log2FC | P | log2FC | P | ||||||||||||||||||||||
| Com_18815_pos | 6-羟基褪黑素 | - | - | - | - | 4.677 | 4.40E-06 | - | - | 2.710 | 0.017 | ||||||||||||||||||||
| Com_1368_pos | 5-甲氧基吲哚乙酸 | - | - | - | - | - | - | 1.203 | 2E-04 | 1.869 | 2.1E-05 | ||||||||||||||||||||
| Com_264_pos | 吲哚 | - | - | - | - | - | - | - | - | 1.777 | 0.022 | ||||||||||||||||||||
| Com_4506_pos | 吲哚-3-乙酰胺 | -1.068 | 0.025 | - | - | - | - | - | - | 1.155 | 0.029 | ||||||||||||||||||||
| Com_3334_neg | 吲哚-3-乙酸 | -3.148 | 3.43E-05 | - | - | -2.216 | 3E-04 | - | - | - | - | ||||||||||||||||||||
| Com_5768_pos | 千里光宁碱 | -0.873 | 0.022 | - | - | 2.224 | 0.015 | - | - | - | - | ||||||||||||||||||||
| Com_16587_pos | 芦丁 | 0.682 | 0.030 | - | - | 2.070 | 0.007 | - | - | - | - | ||||||||||||||||||||
| Com_6517_neg | 短叶松素 | - | - | - | - | 1.494 | 2E-04 | - | - | - | - | ||||||||||||||||||||
| Com_2741_neg | 芹菜素 | - | - | - | - | 1.192 | 0.002 | - | - | - | - | ||||||||||||||||||||
| Com_3581_pos | 芽子碱甲酯 | - | - | - | - | 1.097 | 0.027 | - | - | - | - | ||||||||||||||||||||
| Com_4121_neg | 柚皮素 | - | - | - | - | 1.036 | 0.006 | - | - | - | - | ||||||||||||||||||||
| Com_15315_pos | 绿原酸 | - | - | - | - | -1.039 | 0.007 | - | - | - | - | ||||||||||||||||||||
| Com_1276_pos | 茉莉酸甲酯 | 0.582 | 0.014 | - | - | -1.932 | 1E-04 | - | - | - | - | ||||||||||||||||||||
| Com_1009_neg | 丙酮酸 | -0.269 | 0.039 | -0.846 | 0.001 | - | - | - | - | - | - | ||||||||||||||||||||
| Com_916_neg | 延胡索酸 | -0.673 | 0.043 | -1.309 | 0.001 | - | - | - | - | - | - | ||||||||||||||||||||
| Com_3882_neg | 松柏苷 | - | - | - | - | - | - | 1.579 | 0.006 | - | - | ||||||||||||||||||||
| Com_19926_pos | 异甘草素 | - | - | - | - | - | - | 0.985 | 0.011 | - | - | ||||||||||||||||||||
| Com_8297_neg | 胆红素 | 0.770 | 0.045 | - | - | -1.446 | 0.026 | -0.987 | 0.001 | - | - | ||||||||||||||||||||
| Com_1848_pos | 东莨菪碱 | 2.510 | 3.0E-07 | - | - | -1.482 | 2.2E-05 | -1.597 | 0.001 | - | - | ||||||||||||||||||||
| Com_335_pos | 胡椒酸 | -1.979 | 6.5E-05 | - | - | - | - | - | - | 2.416 | 0.019 | ||||||||||||||||||||
| Com_3212_neg | 左旋多巴 | - | - | - | - | - | - | - | - | 1.966 | 1.6E-06 | ||||||||||||||||||||
| Com_1857_neg | 香草醛 | -0.693 | 0.041 | -1.144 | 4E-04 | - | - | - | - | 1.797 | 0.011 | ||||||||||||||||||||
| Com_5587_neg | 姜黄素 | 1.865 | 0.001 | -1.851 | 0.003 | -2.213 | 0.001 | -2.728 | 3E-05 | -3.065 | 7E-06 | ||||||||||||||||||||
| Com_4159_pos | 脱镁叶绿酸A | - | - | - | - | - | - | 1.478 | 0.003 | 3.682 | 6E-07 | ||||||||||||||||||||
| Com_6775_pos | L-高丝氨酸 | - | - | 3.784 | 2.3E-06 | 3.536 | 3.6E-09 | 2.697 | 1.1E-04 | 3.867 | 2.7E-06 | ||||||||||||||||||||
| Com_2002_pos | 2-氨基乙基膦酸酯 | - | - | 1.411 | 0.009 | 1.048 | 0.037 | 2.115 | 1.0E-04 | 2.638 | 5.4E-05 | ||||||||||||||||||||
| Com_921_pos | N-乙酰鸟氨酸脱酰基酶 | -1.370 | 8.9E-05 | - | - | 1.528 | 1.0E-04 | - | - | 3.149 | 8.0E-04 | ||||||||||||||||||||
| Com_18818_pos | 3-琥珀酰吡啶 | - | - | - | - | 1.191 | 5.0E-04 | 0.854 | 1.2E-04 | 1.484 | 1.0E-06 | ||||||||||||||||||||
| Com_1132_pos | 4-胍基丁酸 | - | - | - | - | - | - | - | - | 3.180 | 0.009 | ||||||||||||||||||||
| Com_205_neg | 棕榈酸 | - | - | - | - | - | - | 0.913 | 0.007 | - | - | ||||||||||||||||||||
| Com_15501_pos | 延胡索碱 | - | - | 1.435 | 3.0E-04 | - | - | - | - | - | - | ||||||||||||||||||||
| Com_5038_neg | 尿黑酸 | - | - | 1.175 | 0.001 | - | - | - | - | - | - | ||||||||||||||||||||
| Com_2413_neg | D-葡萄糖二酸 | -1.317 | 8.5E-05 | - | - | 1.350 | 9.6 E-05 | - | - | - | - | ||||||||||||||||||||
| Com_5408_pos | 泛酰巯基乙胺 | - | - | -0.849 | 0.018 | - | - | -1.087 | 0.001 | -1.491 | 1.2E-06 | ||||||||||||||||||||
| Com_4244_pos | 紫草素 | -1.144 | 4.0E-04 | - | - | - | - | - | - | - | - | ||||||||||||||||||||
| Com_7034_pos | 白藜芦醇 | -1.087 | 7.5E-05 | - | - | - | - | - | - | - | - | ||||||||||||||||||||
| Com_8890_pos | 胆绿素 | -1.041 | 0.011 | - | - | - | - | - | - | - | - | ||||||||||||||||||||
| Com_4560_neg | 紫丁香苷 | - | - | - | - | - | - | - | - | - | - | ||||||||||||||||||||
| Com_12390_pos | 牛磺胆酸 | - | - | - | - | - | - | - | - | - | - | ||||||||||||||||||||
| Com_4167_pos | 血根碱 | - | - | - | - | - | - | - | - | - | - | ||||||||||||||||||||
| Com_3430_pos | 依美汀 | - | - | - | - | - | - | - | - | - | - | ||||||||||||||||||||
“-”表示无显著性差异。
“-”indicates no significant difference.
在低浓度及高浓度复合盐碱胁迫下,吲哚-3-乙酸在Xin-17中始终维持对照水平而未发生变化;低浓度下其在草莜1号中同样未见变化,但高浓度胁迫后却发生显著下调表达(P=3E-04)。对该物质在2个品种中的变化进行比较发现,在对照及低浓度条件下,Xin-17中吲哚-3-乙酸的表达量均显著低于草莜1号。吲哚-3-乙酰胺为吲哚-3-乙酸的上游物质,而吲哚为另一分支产物。盐碱处理后,草莜1号中吲哚-3-乙酰胺和吲哚均维持对照水平;而高浓度胁迫下这2种代谢物在Xin-17中均显著上调表达,并达到草莜1号的表达量水平。对于5-甲氧基吲哚乙酸酯,Xin-17的对照组中表达量则略低于草莜1号,盐碱胁迫后,在Xin-17中表达量上调,但整体表达量在品种间无差异。
复合盐碱胁迫下,燕麦次生代谢通路受到明显影响(图9b)。具体而言,苯丙烷类次生代谢通路中不少代谢物出现差异表达;同时,黄酮和类黄酮类次生代谢通路中也发现有差异表达。比较2个品种间的次生代谢物发现,对照组中多数次生代谢物表达量在品种间无显著差异,如芹菜素、柚皮素、芦丁、香草醛和绿原酸等。也有代谢物既无品种间差异,在受到胁迫后也未发生显著差异表达,表达稳定,如紫丁香苷、牛磺胆酸、血根碱和依美汀等。另有代谢物虽有一定的品种间差异,但受胁迫后均未发现显著的差异表达,如紫草素、白藜芦醇和胆绿素等。此外,L-高丝氨酸和2-氨基乙基膦酸酯作为参与多个代谢途径的代谢物,在受到胁迫后均未发现有显著差异表达;而丙酮酸和延胡索酸在草莜1号中虽有一定的下调,但整体均维持相对稳定水平,品种间均无显著差异。这反映了燕麦响应盐碱胁迫过程中的一些较普遍的响应机制。
当受到高浓度盐碱胁迫后,一些次生代谢物出现明显的品种间差异,这主要由于草莜1号的表达量变化所致,而在Xin-17中多数能稳定表达,与其对照水平无差异。值得注意的是,受到盐碱胁迫后,尽管部分次生代谢物在2个品种内均出现显著下调表达,但Xin-17中的姜黄素和东莨菪碱无论是在对照组还是处理组中,均始终高于在草莜1号中的表达量,特别是东莨菪碱的表达量,品种间最大表现出约10倍(P=1.7E-07)的差异,其次还有胡椒酸和左旋多巴。在受到高浓度盐碱胁迫后,Xin-17中左旋多巴有一定程度的上调表达,但品种间无显著差异,而Xin-17中的松柏甙含量则显著高于草莜1号。
另外,脱镁叶绿酸A作为Chla降解产物,随着盐碱浓度增加,在Xin-17中含量显著增加,从2.8倍(P=0.003)增至12.8倍(P=6E-07),这与该品种中光合色素含量随盐碱浓度升高而急剧下降的结果一致;脱镁叶绿酸A含量在品种间无显著差异(P=0.146),随盐碱浓度升高,在Xin-17中含量逐渐增多,T2处理下Xin-17为草莜1号的2.277倍(P=0.008),T5处理下达到9.569倍(P= 2.9E-04)。
2.5 qRT-PCR结果分析
为更深入解析复合盐碱胁迫对燕麦苗期代谢的影响,选取与KEGG富集代谢通路相关的多个基因,对这些基因的表达量进行荧光定量PCR分析(图10)。其中,YUCCA基因家族的Cluster- 38721.80008(indole-3-pyruvate monooxygenase)基因表达量随着盐碱胁迫浓度升高,在草莜1号中下调表达,而在Xin-17中上调表达,特别是T5处理下该基因的表达量变化显著,这种趋势与草莜1号中的Com_3334_neg(indole-3-acetic acid)表达趋势接近。
Cluster-38721.50882(phenylalanine ammonia- lyase)为苯丙烷类次生代谢途径的重要合成酶基因,在Xin-17中表达量随着胁迫浓度升高,出现先升高后降低趋势,且变化显著,草莜1号中也有类似趋势,但均不显著。高浓度胁迫下Cluster- 38721.47252(sucrose synthase)在Xin-17中表达量显著增加。Cluster-38721.55805(starch synthase)在Xin-17中表达量随胁迫浓度增加呈升高趋势,但不显著,在草莜1号中则表现为先降低后升高的趋势,这与二者干物质的变化趋势一致。同时,KEGG富集后发现Xin-17中糖代谢活跃度高于草莜1号。在根系组织中,随着胁迫浓度的升高,Cluster- 38721.77689(high-affinity nitrate transporter 2.2)的表达量呈现出先显著升高后降低的趋势,Cluster- 38721.84959(phosphate transporter 2)也表现出同样的趋势,但不显著。
图10
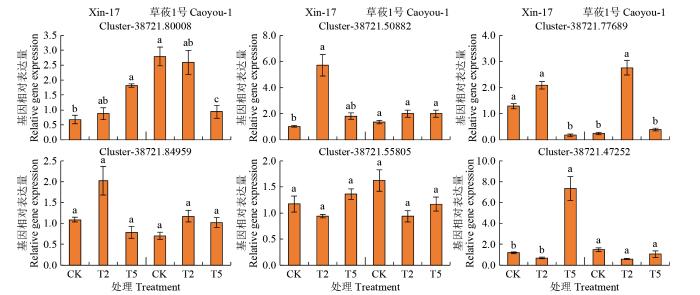
图10
复合盐碱胁迫下6个基因qRT-PCR检测结果
Fig.10
Expression analysis for six genes by qRT-PCR under compound saline-alkali stress
3 讨论
综合表型及生理指标发现,30 mmol/L复合盐碱(T2)对燕麦属于轻度胁迫,而60 mmol/L复合盐碱(T5)产生了显著影响。同时,2个燕麦品种表现出明显的差异,Xin-17的出苗情况及幼苗表型受盐碱胁迫影响均较小。与之相对应的生理指标反映出2个不同抗性品种的不同生理响应特点,抗性品种Xin-17能维持较好的生长发育,这与其保持较高的地上部含水量有关;在受到盐碱胁迫后,Xin-17中光合色素含量开始下调,但仍然维持较高的水平,这将有助于其光合效率的稳定。
生长素在植物的组织发育和细胞伸长等方面发挥着重要作用。在盐碱胁迫下,生长素在物质合成及运输、信号传导等多种生命活动中起到平衡协调作用;同时,其合成通路中的几个关键基因会受盐碱胁迫影响而显著差异表达[27],对应的生长素含量出现显著变化;此外,盐碱胁迫也会影响生长素在植物组织中的分布[28]。本研究中,吲哚-3-乙酸在Xin-17中保持稳定,而草莜1号在T5处理下吲哚-3-乙酸显著下调表达。品种间比较发现,虽然在Xin-17对照组中吲哚-3-乙酸及其上游的吲哚-3-乙酰胺含量均显著低于草莜1号,但是却始终维持稳定表达;而在草莜1号中遭受盐碱胁迫后呈显著下调表达。综上,Xin-17中吲哚-3-乙酸能够维持较低却很稳定的表达,对促进其较高的出苗率和较好的长势有积极作用;尽管Xin-17的长势优于草莜1号是多种因素共同作用的结果,但这种关联仍是探究燕麦耐盐碱机制与耐盐碱品种筛选的一个重要方向,其具体的机制需进一步研究。
植物次生代谢在植物生长调节、传粉、物种竞争和互利共生、适应环境及抵御胁迫过程中均发挥着重要作用[33]。研究[33-34]发现,盐碱胁迫下,植物多酚类物质含量增加,并参与植物细胞中活性氧自由基的清除,酚类物质如黄酮、芦丁、香草醛和绿原酸等[33]会发生显著的差异表达。本研究发现,正常条件下2个品种中次生代谢物的表达变化多数趋于一致,反映了次生代谢在燕麦响应盐碱胁迫中的普遍性;差异则体现在Xin-17中的姜黄素在遭受盐碱胁迫而发生下调表达时,含量仍显著高于草莜1号,较高含量的姜黄素有利于抵御盐碱胁迫。而在低浓度盐碱胁迫下,草莜1号中出现的差异表达代谢物如千里光宁碱上调表达4.65倍,芦丁上调表达约4.2倍,这些增加的次生代谢产物将有助于活性氧自由基的清除,有利于其抵御盐碱胁迫。
4 结论
复合盐碱胁迫下,抗性品种Xin-17的出苗率及苗期的各项指标明显优于草莜1号,这与Xin-17中维持较高的光合色素含量及稳定的地上部含水量,特别是吲哚-3-乙酸的稳定表达,以及姜黄素的较高表达量等结果一致。然而,盐碱胁迫下Xin-17中Chla降解产物脱镁叶绿酸A的显著上调表达,也是该品种的明显缺陷,这将不利于该品种在盐碱胁迫下长期生存。
6-羟基褪黑素在2个品种中均显著上调,这对于燕麦抵御盐碱胁迫具有重要意义。在响应盐碱胁迫过程中,紫丁香苷、牛磺胆酸、血根碱和依美汀等次生代谢物在2个品种中有许多相同的变化趋势,无品种间差异,在受到胁迫后也未发生显著差异表达;也有次生代谢物具有明显的品种特性,如草莜1号中的千里光宁碱和芦丁显著上调,而Xin-17中姜黄素的表达量相对较高。
参考文献
The state of food and agriculture 2021
(2021-12-12)[2024-08-26].https://www.fao.org/3/cb4476en/cb4476en.pdf.
Soil salinization research in China: advances and prospects
Classification of salt-affected soils
Phytoremediation of sodic and saline-sodic soils
Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes
DOI:10.1093/aob/mcu217
PMID:25466549
[本文引用: 1]

Halophytes are the flora of saline soils. They adjust osmotically to soil salinity by accumulating ions and sequestering the vast majority of these (generally Na(+) and Cl(-)) in vacuoles, while in the cytoplasm organic solutes are accumulated to prevent adverse effects on metabolism. At high salinities, however, growth is inhibited. Possible causes are: toxicity to metabolism of Na(+) and/or Cl(-) in the cytoplasm; insufficient osmotic adjustment resulting in reduced net photosynthesis because of stomatal closure; reduced turgor for expansion growth; adverse cellular water relations if ions build up in the apoplast (cell walls) of leaves; diversion of energy needed to maintain solute homeostasis; sub-optimal levels of K(+) (or other mineral nutrients) required for maintaining enzyme activities; possible damage from reactive oxygen species; or changes in hormonal concentrations.This review discusses the evidence for Na(+) and Cl(-) toxicity and the concept of tissue tolerance in relation to halophytes.The data reviewed here suggest that halophytes tolerate cytoplasmic Na(+) and Cl(-) concentrations of 100-200 mm, but whether these ions ever reach toxic concentrations that inhibit metabolism in the cytoplasm or cause death is unknown. Measurements of ion concentrations in the cytosol of various cell types for contrasting species and growth conditions are needed. Future work should also focus on the properties of the tonoplast that enable ion accumulation and prevent ion leakage, such as the special properties of ion transporters and of the lipids that determine membrane permeability.© The Author 2014. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Mechanisms of salinity tolerance
DOI:10.1146/annurev.arplant.59.032607.092911
PMID:18444910
[本文引用: 1]
The physiological and molecular mechanisms of tolerance to osmotic and ionic components of salinity stress are reviewed at the cellular, organ, and whole-plant level. Plant growth responds to salinity in two phases: a rapid, osmotic phase that inhibits growth of young leaves, and a slower, ionic phase that accelerates senescence of mature leaves. Plant adaptations to salinity are of three distinct types: osmotic stress tolerance, Na(+) or Cl() exclusion, and the tolerance of tissue to accumulated Na(+) or Cl(). Our understanding of the role of the HKT gene family in Na(+) exclusion from leaves is increasing, as is the understanding of the molecular bases for many other transport processes at the cellular level. However, we have a limited molecular understanding of the overall control of Na(+) accumulation and of osmotic stress tolerance at the whole-plant level. Molecular genetics and functional genomics provide a new opportunity to synthesize molecular and physiological knowledge to improve the salinity tolerance of plants relevant to food production and environmental sustainability.
Soil salinity: a neglected factor in plant ecology and biogeography
Morphological, physiological, and genetic responses to salt stress in alfalfa: a review
Tuning plant signaling and growth to survive salt
DOI:10.1016/j.tplants.2015.06.008
PMID:26205171
[本文引用: 1]
Salinity is one of the major abiotic factors threatening food security worldwide. Recently, our understanding of early processes underlying salinity tolerance has expanded. In this review, early signaling events, such as phospholipid signaling, calcium ion (Ca(2+)) responses, and reactive oxygen species (ROS) production, together with salt stress-induced abscisic acid (ABA) accumulation, are brought into the context of long-term salt stress-specific responses and alteration of plant growth. Salt-induced quiescent and recovery growth phases rely on modification of cell cycle activity, cell expansion, and cell wall extensibility. The period of initial growth arrest varies among different organs, leading to altered plant morphology. Studying stress-induced changes in growth dynamics can be used for screening to discover novel genes contributing to salt stress tolerance in model species and crops. Copyright © 2015 Elsevier Ltd. All rights reserved.
Plant metabolomics reveals conserved and divergent metabolic responses to salinity
Time-course metabolic profiling in Arabidopsis thaliana cell cultures after salt stress treatment
Ion-dependent metabolic responses of Vicia faba L. to salt stress
Soil processes affecting crop production in salt-affected soils
Transcriptomic and metabolomic analyses reveal mechanisms of adaptation to salinity in which carbon and nitrogen metabolism is altered in sugar beet roots
DOI:10.1186/s12870-020-02349-9
PMID:32245415
[本文引用: 2]
Beta vulgaris L. is one of the main sugar-producing crop species and is highly adaptable to saline soil. This study explored the alterations to the carbon and nitrogen metabolism mechanisms enabling the roots of sugar beet seedlings to adapt to salinity.The ionome, metabolome, and transcriptome of the roots of sugar beet seedlings were evaluated after 1 day (short term) and 7 days (long term) of 300 mM Na treatment. Salt stress caused reactive oxygen species (ROS) damage and ion toxicity in the roots. Interestingly, under salt stress, the increase in the Na/K ratio compared to the control ratio on day 7 was lower than that on day 1 in the roots. The transcriptomic results showed that a large number of differentially expressed genes (DEGs) were enriched in various metabolic pathways. A total of 1279 and 903 DEGs were identified on days 1 and 7, respectively, and were mapped mainly to 10 Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways. Most of the genes were involved in carbon metabolism and amino acid (AA) biosynthesis. Furthermore, metabolomic analysis revealed that sucrose metabolism and the activity of the tricarboxylic acid (TCA) cycle increased in response to salt stress. After 1 day of stress, the content of sucrose decreased, whereas the content of organic acids (OAs) such as L-malic acid and 2-oxoglutaric acid increased. After 7 days of salt stress, nitrogen-containing metabolites such as AAs, betaine, melatonin, and (S)-2-aminobutyric acid increased significantly. In addition, multiomic analysis revealed that the expression of the gene encoding xanthine dehydrogenase (XDH) was upregulated and that the expression of the gene encoding allantoinase (ALN) was significantly downregulated, resulting in a large accumulation of allantoin. Correlation analysis revealed that most genes were significantly related to only allantoin and xanthosine.Our study demonstrated that carbon and nitrogen metabolism was altered in the roots of sugar beet plants under salt stress. Nitrogen metabolism plays a major role in the late stages of salt stress. Allantoin, which is involved in the purine metabolic pathway, may be a key regulator of sugar beet salt tolerance.
Integrated proteomics and metabolomics for dissecting the mechanism of global responses to salt and alkali stress in Suaeda corniculata
Metabolic changes associated with differential salt tolerance in sorghum genotypes
DOI:10.1007/s00425-020-03437-8
PMID:32761417
[本文引用: 1]
Accumulation of specific metabolites, mainly γ-aminobutyric acid, polyamines, and proline, was essential to homeostasis regulation and differential salt tolerance in sorghum genotypes. Salinity is severe abiotic stress that limits plant growth and development in arid and semi-arid regions. Survival to abiotic stresses depends on metabolic and sometimes even morphological adjustments. We measured the growth parameters, water relations, the content of ions (Na, K, Cl), compatible solutes [some free amino acids (FAAs) including γ-aminobutyric acid (GABA) and proline and soluble carbohydrates) and polyamines (PAs), the activity of PAs metabolism enzymes, and metabolomic profile in plants after 14 days of salt stress treatment. These analyses were to evaluate the influence of metabolomic responses of sorghum genotypes exhibiting sensitivity (CSF18) or tolerance (CSF20) to salinity on plant growth. The salinity promoted growth reductions and induced increases in Na and Cl content and decreases in K content. The water status and osmotic potential (Ψ) were reduced by salt stress, but to minimize damage, especially in the CSF20, the osmolytes and PAs contributed to the osmotic adjustment. The results showed that salinity induced an increase in putrescine (Put) in the sensitive genotype. However, it raised spermidine (Spd), spermine (Spm), and cadaverine (Cad) in the tolerant genotype. In addition, the regulation of polyamine oxidase can be related to Spm and GABA biosynthesis. Differential metabolic changes to salt tolerance include metabolites associated with tricarboxylic acid (TCA) cycle intermediates and the metabolisms of sugars, FAAs, and PAs.
Metabolic GWAS-based dissection of genetic bases underlying the diversity of plant metabolism
Metabolomics, a powerful tool for understanding plant abiotic stress
Metabolomics: current application and prospects in crop production
Effects of saline and alkaline stress on germination, seedling growth, and ion balance in wheat
Shotgun proteomic analysis of soybean embryonic axes during germination under salt stress
DOI:10.1002/pmic.201500283
PMID:26969838
Seed imbibition and radicle emergence are generally less affected by salinity in soybean than in other crop plants. In order to unveil the mechanisms underlying this remarkable salt tolerance of soybean at seed germination, a comparative label-free shotgun proteomic analysis of embryonic axes exposed to salinity during germination sensu stricto (GSS) was conducted. The results revealed that the application of 100 and 200 mmol/L NaCl stress was accompanied by significant changes (>2-fold, P<0.05) of 97 and 75 proteins, respectively. Most of these salt-responsive proteins (70%) were classified into three major functional categories: disease/defense response, protein destination and storage and primary metabolism. The involvement of these proteins in salt tolerance of soybean was discussed, and some of them were suggested to be potential salt-tolerant proteins. Furthermore, our results suggest that the cross-protection against aldehydes, oxidative as well as osmotic stress, is the major adaptive response to salinity in soybean.© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Physiological and metabolic responses of Leymus chinensis seedlings to alkali stress
World Oat Production, Trade, and Usage
The role of oat nutrients in the immune system: a narrative review
Phytoremediating coastal saline soils with oats: accumulation and distribution of sodium, potassium, and chloride ions in plant organs
Physiological responses of contrasting rice genotypes to salt stress at reproductive stage
DOI:10.1016/j.rsci.2019.05.001
[本文引用: 1]
Salinity is a major abiotic stress affecting plant growth and productivity. Considerable genetic variation is present in rice in response to salt stress, with higher sensitivity during early seedling and reproductive stage. In this study, physiological changes in leaves and developing panicles of rice genotypes (IR686, Sadri, Rc222, CSR28, IR670 and Pokkali) contrasting in salt tolerance at the reproductive stage were evaluated in greenhouse experiment under salt stress. The results showed that IR670 and the tolerant-check Pokkali maintained lower Na+/K+ ratio, less reduction in chlorophyll concentration, lower malondialdehyde (MDA) production, higher concentrations of reduced ascorbate (reduced AsA), higher proline accumulation and lower percentage reduction in pollen viability than the salt-sensitive genotypes under salt stress. The higher concentration of reduced AsA suggests an efficient ROS-scavenging system. Physiological measurements and pollen viability analysis revealed that Sadri (moderately tolerant at the seedling stage) is sensitive to salt stress at the flowering stage. The findings will be useful in breeding salt tolerant varieties at both seedling and reproductive stages by selecting appropriate genotypes and phenotypes.
Auxin-mediated responses under salt stress: from developmental regulation to biotechnological applications
DOI:10.1093/jxb/eraa241
PMID:32433743
[本文引用: 1]
As sessile organisms, plants are exposed to multiple abiotic stresses commonly found in nature. To survive, plants have developed complex responses that involve genetic, epigenetic, cellular, and morphological modifications. Among different environmental cues, salt stress has emerged as a critical problem contributing to yield losses and marked reductions in crop production. Moreover, as the climate changes, it is expected that salt stress will have a significant impact on crop production in the agroindustry. On a mechanistic level, salt stress is known to be regulated by the crosstalk of many signaling molecules such as phytohormones, with auxin having been described as a key mediator of the process. Auxin plays an important role in plant developmental responses and stress, modulating a complex balance of biosynthesis, transport, and signaling that among other things, finely tune physiological changes in plant architecture and Na+ accumulation. In this review, we describe current knowledge on auxin's role in modulating the salt stress response. We also discuss recent and potential biotechnological approaches to tackling salt stress.© The Author(s) 2020. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Plant hormones in salt stress tolerance
The role of melatonin in salt stress responses
Melatonin: a small molecule but important for salt stress tolerance in plants
Melatonin positively influences the photosynthetic machinery and antioxidant system of Avena sativa during salinity stress
Melatonin and phytomelatonin: chemistry, biosynthesis, metabolism, distribution and bioactivity in plants and animals - an overview

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}