


植物常常通过根系形态变化来应对非生物胁迫,如形成气生组织和不定根以改善根系内的气体交换或增强吸收能力[6-7]。由于根系缺乏通气组织,在各类非生物胁迫中,淹水胁迫对油菜造成的损害最为严重、产量降幅也最大[8-9]。淹水通过阻碍根系吸收养分并破坏根细胞,导致叶片过早老化和光合作用受阻[10]——这主要是由于叶片气孔关闭和叶绿素降解所致。土壤中O2扩散缓慢且供应有限,会促使根系代谢从有氧模式转向无氧模式,进而引发生长迟缓和产量下降[11]。此外,水涝还会影响油料作物的脂质代谢。以花生[12]和向日葵[13]为例,在胁迫条件下,其饱和脂肪酸含量会减少,而不饱和脂肪酸含量则相应增加。油菜籽的品质主要取决于油含量和脂肪酸组成(如油酸等),而水涝胁迫会干扰其脂质代谢,导致品质下降。研究[4]表明,在油菜籽早花期,水涝会增加油酸和硫苷含量,同时降低亚油酸含量。
针对耐渍作物品种培育及耐渍机制解析,此前已进行了诸多研究。Zou等[14]在油菜4~6叶期进行田间淹水胁迫处理,发现油菜耐湿性存在显著的遗传变异,不同甘蓝型油菜品种的耐湿性在产量上存在差异,对涝渍非常敏感的品种产量损失达到50%以上,最耐受的品种产量仅下降10%。Hussain等[15]发现,过量硫可作为调节剂,一定程度上缓解淹水胁迫对油菜的不利影响并促进幼苗生长。Li等[16]发现,BnaPGIP2可通过延缓根细胞壁中果胶的降解,帮助油菜应对淹水胁迫并增强耐渍性。李浩杰等[17]采用室内淹水和田间模拟渍害对甘蓝型油菜耐渍性进行鉴定,发现油菜经胁迫后,根鲜重、根长和株高等指标下降,同时株高、有效角果数和单株产量等发生极显著降低,而可溶性糖、丙二醛(MDA)、脯氨酸(Pro)含量和超氧化物歧化酶(SOD)活性都升高。唐章林等[18]采用盆栽方式在油菜四叶一心时进行渍害胁迫处理,发现胁迫后幼苗绿叶数、地上部鲜质量等性状显著升高。Hare等[19]在塑料桶中对不同叶龄的6种热带草渍水胁迫0、10、20和30 d,发现在淹水胁迫条件下存活并至少保持干重的能力可作为耐湿性鉴定的主要指标。
前人[20]关于甘蓝型油菜耐湿性的鉴定,多选取萌发期、苗期或成熟期进行淹水胁迫处理并鉴定,但涉及多个生育期的综合性鉴定报道较少。萌发期和幼苗期对于作物生长发育而言均十分重要,萌发率的降低将直接减少出苗基数,而幼苗期耐受性不足则易导致弱苗、死苗,二者严重影响群体建成及后续产量形成。基于此,本研究通过2个试验周期的萌发期密闭淹水缺氧处理与苗期大田淹水胁迫处理相结合,并测量淹水胁迫处理后萌发期生理指标、苗期生长状态及后续成熟期农艺性状和品质性状的变化,进而筛选出便捷、易操作的耐湿性鉴定方法,建立较为可靠的油菜耐湿性鉴定评价体系,并据此筛选油菜耐湿种质资源,为遗传改良奠定基础。
1 材料与方法
1.1 供试材料
供试材料为南充市农业科学院油菜研究所种质资源库中经耐湿性预备试验初筛(参试材料60份,调查幼苗死亡率、萎焉系数,并在淹水后进行田间观察),选择耐湿性有明显差异且具有不同遗传背景的21份甘蓝型油菜品种(系),详见表1。
表1 供试材料
Table 1
| 编号 Code | 品种(系) Variety (line) | 编号 Code | 品种(系) Variety (line) |
|---|---|---|---|
| 1 | M417(CV) | 12 | ZS9 |
| 2 | ZS4///南A2/B94076//中双5号 | 13 | ZS6/W757 |
| 3 | F219 | 14 | T8 |
| 4 | T8 | 15 | R896-2 |
| 5 | NY18 | 16 | WY29 |
| 6 | 18C | 17 | ZS4/W757 |
| 7 | Z254 | 18 | FC03(CV) |
| 8 | ZS11 | 19 | 130532-1 |
| 9 | AGREV012 | 20 | 德油早1号 |
| 10 | D57 | 21 | 楚0708(CV) |
| 11 | AGREV012 |
1.2 试验方法
1.2.1 苗期田间淹水处理
试验在南充市农业科学院潆溪科研试验基地(30°52′53″ N,106°2′46″ E)完成,田间试验开展2个周期。前茬为水稻,土质为黄泥土,地势平坦,排灌方便,肥力中等,地力均匀。试验设CK不淹水(T0)、淹水10 d(T1)、淹水15 d(T2)和淹水20 d(T3)4个处理,采用随机区组设计,2次重复,重复间设走道(宽0.6 m)。每份材料种植面积1.2 m2,2行区,每行10株,直播规格50 cm×12 cm。选择在幼苗第6~7片真叶长出时,进行淹水处理,各淹水处理间相距1.5 m,为防止不同淹水处理之间相互窜水,不同淹水处理间垒土坝加固,在各处理厢面四周开深沟,铺埋塑料膜设置隔水层。用加厚塑料膜覆盖土坝两侧并密封固定,与周围隔离,不同处理间开0.6 m深沟铺埋塑料薄膜隔水。淹水期间保持水层在厢面3 cm左右,淹水结束后迅速去水解除胁迫。
1.2.2 苗期生长性状
在达到预定淹水处理天数后,排干水正常生长7 d,各处理选取每小区第2行连续5株,分别统计和测量茎上所有的绿叶数和苗高,按照农业气象相关规范,能进行光合作用的叶片均为绿叶,没展开的新叶也一并计入,部分黄化的叶片则按绿色部分占整个叶片的百分比进行计算,如绿色部分面积占整个叶片面积的30%,其余部分已变黄,则将其记为0.3片绿叶,此为目测估算值,结果保留1位小数;苗高为从子叶节到顶端分生组织(生长点)的长度。
1.2.3 成熟期农艺性状
成熟期每份材料随机取5株测量考种,3次重复,涉及株高、一次有效分枝高度、一次有效分枝数、主花序长度、主花序有效角果数、主花絮无效角果数、一分枝有效角果数、单株有效角果数、每果粒数、千粒重和经济产量等。株高指从子叶节到整株油菜主茎最高点的长度;一次有效分枝高度是指子叶节与主茎上最下部第一个有效分枝着生点之间的距离;一次有效分枝数指直接从主茎上长出的有一个有效角果以上的分枝数目;经济产量是指5株植株所有自然风干种子总重量的平均值。
1.2.4 品质性状
每份材料随机取5株单独脱粒并进行3次重复,采用近红外光谱法测定油菜籽芥酸、硫苷、含油量、蛋白质、亚麻酸、亚油酸和花生烯酸含量。
1.2.5 萌发期生理指标
通过2个周期大田淹水试验,从21份材料中鉴别出耐湿性强弱差异显著的12份材料进行试验,参考并适当改进Langan等[20]的方法,选取饱满种子置于滤纸发芽床中,于25 ℃萌发60 h,取露白后48 h的芽苗(胚根长2~3 mm)50粒转移到50 mL带盖PVC离心管中,无菌淹水缺氧处理24 h(24H),用无菌水冲洗3~4次后,置于1 cm左右厚湿润细砂(细砂预先于105 ℃的烘箱处理24 h)发芽床中,再覆盖一层细砂,保持湿润,于25 ℃继续发芽6 d,以不经水淹处理的芽苗为对照(CK),CK的露白种子不作水淹处理直接进行沙培。6 d后,24H和CK处理每份材料随机选取4~5株苗准确称量0.25 g,混合磨样,采用南京建成生物工程研究所研发的蛋白定量(TP)测定试剂盒(A045-2考马斯亮蓝法)、MDA测定试剂盒(A003-1 TBA法)、过氧化物酶(POD)测试试剂盒(A084-3-1比色法)、Pro测定试剂盒(A107-1-1比色法)、植物铜锌―超氧化物歧化酶(Cu2+-Zn2+-SOD)测定试剂盒(A001-4-1比色法)分别测定可溶性蛋白(soluble protein,SP)、MDA、Pro含量及POD、植物铜锌―超氧化物歧化酶(Copper-zinc superoxide dismutase,Cu-Zn SOD)和SOD活性。
1.3 数据处理
2 结果与分析
2.1 苗期淹水处理前绿叶数与苗高
由表2可知,淹水处理、淹水处理与材料互作间的绿叶数、苗高差异不显著,表明淹水处理前各处理间苗情一致,绿叶数和苗高2个性状在不同材料间均存在极显著差异,表明供试材料本身存在遗传差异,符合试验设计要求,可进行下一步淹水处理试验。
表2 21份油菜品种(系)淹水前绿叶数、苗高的方差分析
Table 2
| 变异来源 Source of variation | 自由度 df | F值F-value | |
|---|---|---|---|
| 绿叶数 Number of green leaves | 苗高 Seedling height | ||
| 淹水处理Flooding treatment | 3 | 2.302 | 1.853 |
| 材料Material | 20 | 2.411** | 3.894** |
| 淹水处理×材料 Flooding treatment×material | 60 | 0.310 | 0.522 |
| 误差Error | 84 | ||
“**”表示差异达P < 0.01显著水平,下同。
“**”indicates significant difference at P < 0.01 level, the same below.
2.2 苗期淹水处理后绿叶数与苗高
表3 21份油菜品种(系)淹水后绿叶数、苗高的方差分析
Table 3
| 变异来源 Source of variation | 自由度 df | F值F-value | |
|---|---|---|---|
| 绿叶数 Number of green leaves | 苗高 Seedling height | ||
| 淹水处理Flooding treatment | 2 | 9.974** | 12.939** |
| 材料Material | 20 | 2.299** | 2.731** |
| 淹水处理×材料 Flooding treatment×material | 40 | 0.542 | 0.414 |
| 误差Error | 63 | ||
表4 淹水处理绿叶数、苗高与CK差值均值成对比较
Table 4
| 处理 Treatment | 绿叶数Number of green leaves | 苗高Seedling height | |||||
|---|---|---|---|---|---|---|---|
| (对照-处理)均值 (Control-treatment) mean | 处理 Treatment | 均值差 Mean difference | (对照-处理)均值 (Control-treatment) mean | 处理 Treatment | 均值差 Mean difference | ||
| T3 | 0.919 | T3-T2 | 0.516* | 1.739 | T3-T2 | 1.748** | |
| T2 | 0.406 | T3-T1 | 0.910** | -0.009 | T3-T1 | 2.510** | |
| T1 | 0.010 | T2-T1 | 0.397* | -0.771 | T2-T1 | 0.761 | |
“*”表示差异达P < 0.05显著水平,下同。
“*”indicates significant difference at P < 0.05 level, the same below.
2.3 淹水处理后农艺性状及产量性状
表5 21份油菜品种(系)2个试验周期成熟期农艺性状
Table 5
| 处理 Treatment | 试验周期 Test cycle | 株高 Plant height (cm) | 一次有效分枝高度 Primary effective branch height (cm) | 主花序长度 Main inflorescence length (cm) | 一次有效分枝数 Number of primary effective branches | 主花序无效角果数 Invalid pods in main inflorescence |
|---|---|---|---|---|---|---|
| T0 (CK) | 1 | 73.503±20.542 | 51.658±12.004 | 20.636±9.730 | 0.379±0.823 | 3.800±3.536 |
| 2 | 152.784±25.845 | 54.730±19.472 | 57.681±8.932 | 6.410±1.140 | 7.863±5.684 | |
| T1 | 1 | 80.952±14.288 | 54.098±10.769 | 24.771±7.655 | 0.467±0.795 | 3.993±2.576 |
| 2 | 142.906±27.475 | 49.815±19.225 | 55.291±9.951 | 5.972±1.523 | 6.061±5.688 | |
| T2 | 1 | 70.279±17.362 | 50.153±12.198 | 19.729±7.158 | 0.139±0.228 | 4.023±2.728 |
| 2 | 138.369±26.673 | 43.163±16.680 | 56.008±10.628 | 6.060±1.313 | 6.357±5.190 | |
| T3 | 1 | 67.752±20.096 | 46.324±11.371 | 21.404±11.801 | 0.251±0.552 | 4.436±3.631 |
| 2 | 128.920±22.014 | 36.526±15.216 | 53.220±10.775 | 5.954±1.676 | 6.832±4.467 | |
| 处理 Treatment | 试验周期 Test cycle | 主花序有效角果数 Effective pods in main inflorescence | 一分枝有效角果数 Number of pods per primary branch | 每果粒数 Number of seeds per pod | 单株角果数 Number of pods per plant | |
| T0 (CK) | 1 | 19.314±9.457 | 2.478±6.932 | 13.816±4.069 | 21.019±14.361 | |
| 2 | 68.932±19.045 | 257.756±105.979 | 13.727±5.971 | 326.687±14.361 | ||
| T1 | 1 | 23.152±6.392 | 3.117±6.617 | 16.313±2.556 | 26.195±11.141 | |
| 2 | 64.548±16.998 | 202.609±85.321 | 13.924±5.413 | 299.119±113.963 | ||
| T2 | 1 | 19.685±7.730 | 0.563±1.244 | 13.973±3.750 | 20.248±8.425 | |
| 2 | 58.516±15.245 | 248.655±119.895 | 11.919±5.370 | 307.171±129.735 | ||
| T3 | 1 | 18.704±8.881 | 2.110±5.323 | 14.402±3.403 | 20.263±12.378 | |
| 2 | 51.271±19.225 | 247.254±168.329 | 12.822±7.727 | 20.281±12.439 | ||
表6 21份油菜品种(系)经济产量及千粒重
Table 6
| 处理 Treatment | 经济产量(g/株) Economic yield (g/plant) | 千粒重 1000-grain weight (g) |
|---|---|---|
| T0 (CK) | 149.885±68.782 | 3.491±0.569 |
| T1 | 143.129±61.904 | 3.569±0.561 |
| T2 | 114.744±51.910 | 3.293±0.520 |
| T3 | 107.316±47.155 | 3.376±0.606 |
2.4 淹水处理后品质性状
表7 21份油菜品种(系)的品质性状
Table 7
| 处理 Treatment | 试验周期 Test cycle | 芥酸 Erucic acid | 硫苷 Sulfur glycoside | 含油量 Oil content | 蛋白质含量 Protein content | 亚麻酸 Linolenic acid | 亚油酸 Linoleic acid | 花生烯酸 Arachidonic acid |
|---|---|---|---|---|---|---|---|---|
| T0 | 1 | 0.294±0.580 | 36.554±12.045 | 43.779±3.373 | 17.753±2.531 | 8.810±2.107 | 18.035±1.908 | 3.676±2.135 |
| 2 | 0.738±0.423 | 29.032±6.202 | 42.896±3.977 | 24.193±2.175 | 10.022±1.194 | 16.843±1.948 | 0.566±1.125 | |
| T1 | 1 | 0.310±0.529 | 31.125±11.523 | 42.414±3.458 | 18.939±2.201 | 10.258±1.501 | 18.896±1.969 | 2.301±1.185 |
| 2 | 0.548±0.380 | 28.026±6.528 | 43.560±2.987 | 23.138±2.041 | 9.874±0.941 | 15.957±1.784 | 0.100±1.234 | |
| T2 | 1 | 0.232±0.455 | 33.913±11.791 | 43.675±3.299 | 18.257±2.097 | 9.391±1.987 | 18.446±1.871 | 2.894±1.617 |
| 2 | 0.694±0.378 | 29.883±7.206 | 42.326±3.047 | 23.707±2.259 | 9.844±1.035 | 16.794±1.822 | 0.141±1.092 | |
| T3 | 1 | 0.243±0.529 | 35.553±12.446 | 43.554±2.968 | 18.179±1.834 | 8.535±2.173 | 18.290±1.699 | 3.196±1.642 |
| 2 | 0.620±0.360 | 27.915±5.861 | 41.799±4.592 | 23.999±2.236 | 9.906±1.299 | 16.793±2.123 | 0.041±1.145 |
表8 2个试验周期21份油菜品种(系)品质性状的方差分析
Table 8
| 变异来源 Source of variation | 自由度 df | 试验 周期 Test cycle | F值F-value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 芥酸 Erucic acid | 硫苷 Sulfur glycoside | 含油量 Oil content | 蛋白质含量 Protein content | 亚麻酸 Linolenic acid | 亚油酸 Linoleic acid | 花生烯酸 Arachidonic acid | |||
| 淹水处理 Flooding treatment | 3 | 1 | 0.754 | 2.257 | 7.997** | 8.248** | 8.489** | 5.565** | 8.386** |
| 2 | 3.763* | 2.936* | 4.161** | 4.491** | 0.280 | 8.748** | 4.427** | ||
| 材料 Material | 20 | 1 | 1.914** | 4.381** | 33.253** | 22.688** | 4.118** | 21.264** | 6.290** |
| 2 | 7.665** | 14.733** | 11.813** | 14.011** | 15.581** | 33.059** | 17.935** | ||
| 淹水处理×材料 Flooding treatment×material | 60 | 1 | 0.890 | 0.871 | 1.415 | 1.620* | 0.883 | 1.127 | 1.143 |
| 2 | 2.392** | 1.971** | 2.319** | 2.390** | 2.019** | 3.640** | 1.470 | ||
| 误差Error | 84 | ||||||||
2.5 萌发期淹水处理后生理指标测定
萌发初期的种子经过24 h淹水处理后,在正常条件下培养6 d对相关指标进行测定。如表9所示,与CK相比,处理后SPC、MDA、Pro、POD和总SOD(T-SOD)活性均有不同程度升高,尤其是MDA含量升高明显。
表9 12份油菜品种(系)萌发期淹水胁迫下生理指标测定
Table 9
| 处理 Treatment | 可溶性蛋白 SP (μg/g) | 丙二醛 MDA (nmol/mg prot) | 脯氨酸 Pro (U/g) | 过氧化物酶 POD (U/g) | 铜锌超氧化物歧化酶 Cu-Zn SOD (U/g) | 超氧化物歧化酶 T-SOD (U/g) |
|---|---|---|---|---|---|---|
| CK | 1.732±0.439 | 0.132±0.055 | 1.062±0.718 | 575.103±136.912 | 645.564±80.982 | 843.571±91.819 |
| 24H | 1.836±0.541 | 0.340±0.100 | 1.267±0.819 | 609.713±167.402 | 647.650±59.027 | 866.106±82.568 |
2.6 主成分分析
采用从农艺性状、产量性状和品质性状等中选取在淹水处理间和材料间都存在极显著差异的11个性状的耐湿系数进行主成分分析,各主成分因子特征值、贡献率和累计贡献率见表10。前7个因子累计贡献率为94.57%,可代表所有性状的全部信息。
表10 主成分因子特征值、贡献率及累计贡献率
Table 10
| 主成分因子 Principal component factor | 特征值 Eigenvalue | 贡献率 Contribution rate (%) | 累计贡献率 Cumulative contribution rate (%) | 主成分因子 Principal component factor | 特征值 Eigenvalue | 贡献率 Contribution rate (%) | 累计贡献率 Cumulative contribution rate (%) |
|---|---|---|---|---|---|---|---|
| PC1 | 3.90 | 35.42 | 35.42 | PC7 | 0.46 | 4.21 | 94.57 |
| PC2 | 2.05 | 18.67 | 54.09 | PC8 | 0.28 | 2.56 | 97.12 |
| PC3 | 1.64 | 14.88 | 68.98 | PC9 | 0.17 | 1.51 | 98.63 |
| PC4 | 1.11 | 10.06 | 79.04 | PC10 | 0.10 | 0.94 | 99.57 |
| PC5 | 0.65 | 5.95 | 84.99 | PC11 | 0.05 | 0.43 | 100.00 |
| PC6 | 0.59 | 5.37 | 90.36 |
由表11可知,PC1中经济产量、绿叶数、含油量、株高和每果粒数占比较大;PC2中苗高、株高、绿叶数和蛋白质含量占比较大;PC3中一次有效分枝数及亚油酸、含油量、千粒重和蛋白质含量占比较大;PC4中一次有效分枝数、株高、亚油酸含量和每果粒数占比较大;PC5中亚油酸含量和株高占比较大;PC6中,蛋白质含量、绿叶数、苗高、花生烯酸含量、经济产量和亚油酸占比较大;PC7中苗高、花生烯酸、亚油酸和含油量占比较大。
表11 主成分因子特征向量值
Table 11
| 性状 Trait | 主成分因子Principal component factor | ||||||
|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | |
| 绿叶数Number of green leaves | 0.3929 | 0.2128 | -0.0759 | -0.1498 | 0.0844 | 0.4908 | -0.1435 |
| 苗高Seedling height | 0.2332 | 0.4287 | -0.2843 | 0.0364 | 0.0457 | 0.2717 | 0.6460 |
| 株高Plant height | 0.3737 | 0.2555 | 0.0860 | 0.2959 | 0.2839 | -0.3418 | -0.1828 |
| 一次有效分枝数Number of primary effective branches | 0.1794 | 0.2095 | 0.5243 | 0.3526 | -0.5204 | 0.0418 | -0.1179 |
| 每果粒数Number of seeds per pod | 0.2907 | -0.4206 | -0.2628 | 0.2647 | 0.0237 | 0.0953 | -0.2795 |
| 含油量Oil content | 0.3874 | 0.0649 | 0.3521 | -0.0780 | -0.0237 | -0.3491 | 0.2855 |
| 蛋白质含量Protein content | -0.3353 | 0.2889 | 0.3061 | 0.1071 | 0.0176 | 0.5179 | -0.2268 |
| 亚油酸Linoleic acid | -0.0636 | -0.3242 | 0.4603 | 0.2619 | 0.6509 | 0.2025 | 0.2899 |
| 花生烯酸Arachidonic acid | 0.1817 | -0.5294 | 0.1504 | -0.1017 | -0.4251 | 0.2594 | 0.3360 |
| 经济产量Economic yield | 0.4578 | -0.1079 | -0.0810 | 0.0537 | 0.0995 | 0.2434 | -0.2684 |
| 千粒重1000-grain weight | 0.1604 | 0.0141 | 0.3215 | -0.7713 | 0.1537 | -0.0031 | -0.1864 |
2.7 隶属函数分析
计算11个性状隶属函数值,如表12所示,平均隶属值排名前7位的材料有8、11、15、14、17、5和19。农艺性状的平均隶属函数值排名前7位的材料有15、14、8、17、11、5和10;品质性状的平均隶属函数值排名前7位的材料有11、18、2、17、6、5和8;产量性状的平均隶属函数值排名前7位的材料有19、8、15、11、14、5和4。其中材料8、11、17和5的隶属函数值在农艺性状、品质性状、产量性状中均居于前列,表明其具有较为稳定的耐湿能力,材料2、3、1和20的隶属函数值在农艺性状、品质性状、产量性状中均居于后列,表明耐湿能力较弱。通过计算各材料的主成分因子综合得分,17、11、14、15、8、5和18位列前7。
表12 各性状隶属函数值、主成分因子综合值
Table 12
| 编号 Code | 隶属函数分析MF analysis | 主成分分析PCA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 平均隶属值 Average membership value | 排名 Ranking | 农艺性状平均 隶属函数值 Agronomic trait average membership value | 排名 Ranking | 品质性状平均 隶属函数值 Quality trait average membership value | 排名 Ranking | 产量性状平均 隶属函数值 Yield trait average membership value | 排名 Ranking | 主成分因子 综合值 Comprehensive value of PCA | 排名 Ranking | ||
| 8 | 0.701 | 1 | 0.804 | 3 | 0.550 | 7 | 0.743 | 2 | 2.825 | 5 | |
| 11 | 0.650 | 2 | 0.579 | 5 | 0.779 | 1 | 0.569 | 4 | 5.228 | 2 | |
| 15 | 0.648 | 3 | 0.867 | 1 | 0.403 | 18 | 0.593 | 3 | 4.338 | 4 | |
| 14 | 0.637 | 4 | 0.830 | 2 | 0.469 | 16 | 0.492 | 5 | 4.991 | 3 | |
| 17 | 0.596 | 5 | 0.649 | 4 | 0.629 | 4 | 0.399 | 9 | 6.882 | 1 | |
| 5 | 0.517 | 6 | 0.503 | 6 | 0.575 | 6 | 0.435 | 6 | 2.285 | 6 | |
| 19 | 0.476 | 7 | 0.312 | 17 | 0.538 | 8 | 0.763 | 1 | -2.560 | 18 | |
| 18 | 0.463 | 8 | 0.353 | 15 | 0.743 | 2 | 0.179 | 19 | 1.891 | 7 | |
| 7 | 0.463 | 9 | 0.445 | 9 | 0.513 | 11 | 0.409 | 8 | -0.519 | 9 | |
| 10 | 0.454 | 10 | 0.462 | 7 | 0.526 | 9 | 0.290 | 13 | -0.647 | 11 | |
| 4 | 0.438 | 11 | 0.391 | 13 | 0.511 | 12 | 0.409 | 7 | 0.551 | 8 | |
| 12 | 0.429 | 12 | 0.423 | 10 | 0.524 | 10 | 0.252 | 17 | -2.112 | 15 | |
| 21 | 0.418 | 13 | 0.410 | 11 | 0.496 | 13 | 0.284 | 15 | -2.177 | 16 | |
| 6 | 0.412 | 14 | 0.306 | 18 | 0.590 | 5 | 0.323 | 12 | -1.149 | 12 | |
| 16 | 0.405 | 15 | 0.447 | 8 | 0.414 | 17 | 0.284 | 14 | -1.387 | 14 | |
| 2 | 0.402 | 16 | 0.269 | 20 | 0.655 | 3 | 0.227 | 18 | -2.476 | 17 | |
| 3 | 0.392 | 17 | 0.407 | 12 | 0.493 | 14 | 0.153 | 20 | -0.631 | 10 | |
| 13 | 0.352 | 18 | 0.238 | 21 | 0.477 | 15 | 0.386 | 10 | -5.498 | 21 | |
| 9 | 0.318 | 19 | 0.377 | 14 | 0.242 | 20 | 0.323 | 11 | -4.458 | 20 | |
| 1 | 0.300 | 20 | 0.287 | 19 | 0.335 | 19 | 0.265 | 16 | -4.227 | 19 | |
| 20 | 0.249 | 21 | 0.315 | 16 | 0.230 | 21 | 0.119 | 21 | -1.150 | 13 | |
2.8 相关性分析
通过对萌发期生理指标、苗期生长指标、成熟期产量及品质相关性状的耐湿系数进行相关分析,结果如图1所示,POD活性与经济产量呈正相关,相关系数为0.72;Cu-Zn SOD与经济产量呈正相关,相关系数为0.63。结果表明,通过测定甘蓝型油菜发芽期缺氧淹水处理的POD和Cu-Zn SOD值,可以对各材料的耐湿性进行快速鉴定。
图1
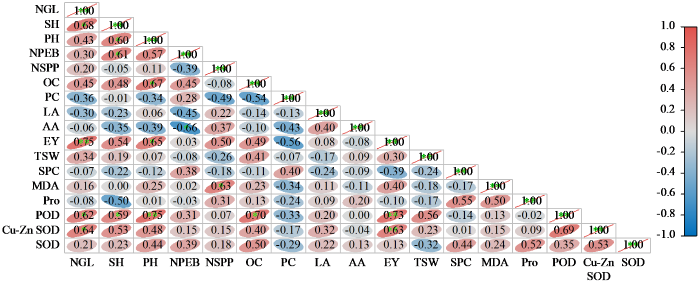
图1
各性状间WTC相关性分析
PC:蛋白质含量;LA:亚油酸;AA:花生烯酸;NSPP:每果粒数;EY:经济产量;TSW:千粒重;NGL:绿叶数;SH:苗高;PH:株高;OC:含油量;NEB:一次有效分枝数;SPC:可溶性蛋白含量。
Fig.1
Correlation analysis of WTC for traits
PC: protein content; LA: linoleic acid; AA: arachidonic acid; NSPP: number of seeds per pod; EY: economic yield; TSW: 1000-grain weight; NGL: number of green leaves; SH: seedling height; PH: plant height; OC: oil content; NPEB: number of primary effective branches; SPC: soluble protein content.
3 讨论
作物耐渍性筛选是鉴定耐渍种质资源的重要手段,通常涉及生理指标、生长发育等多个方面的指标。种子萌发及幼苗期是植物生长发育的初级阶段,也是对渍害最为敏感的时期,该阶段的耐渍性在植株后期生长过程中扮演了十分重要的角色。
3.1 淹水胁迫对表型性状的影响
淹水胁迫对油菜种子产生多方面影响,最直接且显著的表现是表型变化,这种影响几乎贯穿整个生长周期[24]。不同生长阶段的受胁迫程度存在差异,其中以幼苗期最为显著。作为光合作用的主要场所,叶片是植物生长发育的重要能量来源,渍害影响下其碳水化合物代谢和脂质糖化能力受到抑制[25]、绿叶数受到严重影响,这表明淹水胁迫能够严重损害油菜的光合作用并进一步导致发育进程延缓[26-
3.2 淹水胁迫对产量性状的影响
油菜产量与多个性状密切相关,包括每果粒数、一次有效分枝数、主枝角果数以及千粒重等[33],本研究对上述性状均进行了测量,发现几乎所有指标均呈现下降趋势,表明淹水胁迫对产量产生了明显影响,后续对经济产量的测量也证实了这一结论。这些结果与前人[34-35]研究的结论一致,均表明了淹水胁迫下产量与这些性状存在相关性。除产量外,种子质量也是成熟阶段的评估指标之一,包括含油量、脂肪酸组成等。开花期短期淹水胁迫对含油量影响甚微,但若持续时间过长,将严重影响含油量并增加种子中芥酸和硫苷的含量。本研究中上述物质含量的变化趋势与前人研究一致。淹水胁迫可能通过干扰油菜的营养生长来阻碍光合作用和生殖生长进程,最终导致种子发育异常,从而影响油料品质。
3.3 淹水胁迫对萌发期生理特性的影响
呼吸模式的变化可能对生物体的生长发育产生显著影响。在淹水胁迫下的低氧环境中,由于根系进入厌氧模式,加速乙醇积累并形成胞质酸性环境,最终引发氧化爆发并导致萌发率降低、个体生长迟缓。作为氧化爆发的主要产物,活性氧(ROS)的过量生成打破了其积累与清除的平衡,导致生物分子氧化损伤,进而引发细胞损伤和死亡[36-37]。脂质过氧化不仅会破坏DNA和蛋白质功能,还会生成MDA等异常醛类物质[38],这些物质可作为ROS引发的细胞膜损伤程度指标[39-40]。为应对这种情况,抗氧化酶活性和Pro等渗透调节物质会被激活,以维持细胞膨压并减少损伤。本研究通过分析抗氧化酶活性、Pro及MDA含量等指标发现,淹水胁迫显著提升了抗氧化酶活性和脂质过氧化产物含量,且不同品种间存在差异。其中,Cu-ZnSOD在组内和组间均表现出极显著差异,这与前人研究结果一致。综上所述,抗氧化酶系统的增强有助于维持细胞氧化平衡,提升对淹水胁迫压力的耐受性。因此,Cu-Zn SOD和POD可作为抗逆性的重要指标。
4 结论
本研究发现通过测量甘蓝型油菜萌发期的Cu-Zn SOD和POD活性、幼苗期处理20 d后的绿叶数和苗高、成熟期的株高和每果粒数等指标,可有效评估油菜品种的耐渍能力;并利用构建的甘蓝型油菜耐湿性综合评价体系,从21份参试材料中筛选出4个具有较强耐渍性的甘蓝型油菜品种(系)NY18、ZS11、AGREV012和ZS4/W757,以及2个耐渍性较弱的品种(系)M417(CV)和ZS4///南A2/B94076//中双5号。
参考文献
Effect of mineral sulphur availability on nitrogen and sulphur uptake and remobilization during the vegetative growth of Brassica napus L
DOI:10.1093/jxb/erq096
PMID:20403880
[本文引用: 1]

Because it has a high demand for sulphur (S), oilseed rape is particularly sensitive to S limitation. However, the physiological effects of S limitation remain unclear, especially during the rosette stage. For this reason a study was conducted to determine the effects of mineral S limitation on nitrogen (N) and S uptake and remobilization during vegetative growth of oilseed rape at both the whole-plant and leaf rank level for plants grown during 35 d with 300 microM (34)SO(4)(2-) (control plants; +S) or with 15 microM (34)SO(4)(2-) (S-limited plants; -S). The results highlight that S-limited plants showed no significant differences either in whole-plant and leaf biomass or in N uptake, when compared with control plants. However, total S and (34)S (i.e. deriving from S uptake) contents were greatly reduced for the whole plant and leaf after 35 d, and a greater redistribution of endogenous S from leaves to the benefit of roots was observed. The relative expression of tonoplast and plasmalemma sulphate transporters was also strongly induced in the roots. In conclusion, although S-limited plants had 20 times less mineral S than control plants, their development remained surprisingly unchanged. During S limitation, oilseed rape is able to recycle endogenous S compounds (mostly sulphate) from leaves to roots. However, this physiological adaptation may be effective only over a short time scale (i.e. vegetative growth).
Advances of oilseed rape breeding
Responses of plant development, biomass and seed production of direct sown oilseed rape (Brassica napus) to nitrogen application at different stages in Yangtze River Basin
DOI:10.1016/j.fcr.2016.04.024 URL [本文引用: 1]
The effect of waterlogging on yield and seed quality at the early flowering stage in Brassica napus L
DOI:10.1016/j.fcr.2015.06.007 URL [本文引用: 2]
A comparison study of precipitation in the poyang and the dongting lake basins from 1960-2015
DOI:10.1038/s41598-020-60243-8
PMID:32099049
[本文引用: 1]
The Dongting Lake Basin and the Poyang Lake Basin, both located in the middle reaches of the Yangtze River, provide 30% of the total water volume for the Yangtze River. Under global climate change, precipitation patterns have undergone varying degrees of changes in different regions. Analysing temporal and spatial rainfall variations is important for understanding the variations in capacity of the two lake basins and the water intake variations by the Yangtze River. This study analyses the temporal and spatial variations of the two basins based on 33 rain-gauge data series from 1960-2015, using statistical methods, GIS spatial analysis and the M-K trend test. Our results showed that the annual precipitation generally increased in the Poyang Lake Basin and we found no obvious changes in the Dongting Lake Basin from 1960 to 2015. Seasonal precipitation levels at interannual scales were roughly consistent, but exhibited variability larger by an order of magnitude in the Poyang Lake Basin than in the Dongting Lake Basin. In general, an increasing trend dominated in spring and autumn while a decreasing trend was observed in summer and winter. The increasing trend was significant from the 1990s in the Poyang Lake Basin and from the late 1990s in the Dongting Lake Basin. It was found that annual precipitation with relatively larger anomalies appeared in ENSO (El Niño and Southern Oscillation) years in most cases, such as in 1963, 1997/1998 and 2002, while a few anomalies appeared in the previous or next year around an ENSO year, such as 1971 and 1978. All monthly precipitation periods with relatively larger or smaller anomalies coincided with ENSO events. In addition, El Niño and SOI (Southern Oscillation) events had significant relationships with negative monthly precipitation anomalies. El Niño and the SOI exerted more significant impacts on the Poyang Lake Basin than on the Dongting Lake Basin, which explains the conclusions regarding seasonal precipitation trends as mentioned above.
Effects of supplemental nitrogen application on physiological characteristics, dry matter and nitrogen accumulation of winter rapeseed (Brassica napus L.) under waterlogging stress
DOI:10.1038/s41598-020-67260-7
[本文引用: 1]
Waterlogging stress is a common limiting factor for winter rapeseed, which greatly affects the growth and potential production. The present study was conducted to investigate the effects of waterlogging with different durations (0day (D0), 6days (D6) and 9days (D9)) and supplemental nitrogen fertilization (N1, 0 kg ha−1; N2, 30 kg ha−1; N3, 60 kg ha−1 and N4, 90 kg ha−1) on the physiological characteristics, dry matter and nitrogen accumulation in winter rapeseed (Chuanyou36). The results showed that the supplementary application of nitrogen fertilizer could effectively improve the physiological indexes of winter rapeseed in both pot and field experiments. The supplemental nitrogen increased the chlorophyll content in leaves, enhanced the activities of SOD, CAT, and POD, and decreased the MDA content in leaves and roots of rapeseed. The chlorophyll contents, the antioxidant enzyme activity of leaves and roots significantly increased under D6N3 and D9N4 conditions in both (pot and field) experiments. However, MDA contents significantly decreased compared with waterlogging without nitrogen application. Moreover, the application of nitrogen fertilizer after waterlogging increased the accumulation of dry matter and nitrogen in rapeseed at different growth stages. Therefore, waterlogging stress significantly inhibited the growth and development of rapeseed, but the application of nitrogen fertilizer could effectively reduce the damage of waterlogging. The N-induced increase in waterlogging tolerance of rapeseed might be attributed to the strong antioxidant defense system, maintenance of photosynthetic pigments and the nutrient balance.
First report of Sclerotinia sclerotiorum causing sclerotinia blight on peanut (Arachis hypogaea) in Northeastern China
Synergistic effects of plant growth promoting rhizobacteria and silicon dioxide nano-particles for amelioration of drought stress in wheat
DOI:10.1016/j.plaphy.2021.05.039
PMID:34116336
[本文引用: 1]
Drought tolerant plant growth-promoting rhizobacteria (PGPR) can confer drought tolerance in plants, when inoculated, and this effect can be more pronounced by their combined application with silicon oxide nanoparticles (SiO NPs). In this research, drought-tolerant and plant growth-promoting rhizobacterial strains were isolated from the rhizospheric soil of wheat plants growing in the arid region of Pakistan. Out of 30 isolated strains, three rhizobacterial strains were selected based on their drought tolerance, higher phytohormones (indole acetic acid (IAA), abscisic acid (ABA), and cytokinin (CK), and osmolyte (proline and sugar) production ability. These strains were identified as Bacillus sp. Azospirillum lipoferum and Azospirillum brasilense by 16S rRNA sequencing and accession numbers (MT482404, MT742664, and MT 742666, respectively) were obtained. Inoculation of these strains, alone and in combination, improved the germination attributes of wheat seeds under drought stress conditions. However, the combination of all three bacterial strains gave the best results. SiO NPs were prepared from silicon dioxide and characterized by scanning electron microscopy (SEM), Energy dispersive X-rays pattern (EDX), and UV-visible spectrum. The effect of SiO NPs was also tested on wheat seeds under drought stress and it was observed that SiO NPs (150 mg/L) create pronounced drought ameliorative potential in wheat seedlings. In the pot experiment, the combined application of SiO NPs and PGPR exhibited a synergistic role and improved the growth and yield of wheat. The interaction between SiO NPs and bacterial combination improved biomass (fresh and dry weight), and chlorophyll-a, b content by 138.78%, 65.70%, 128.57%, and 283.33% respectively as compared to untreated but drought exposed plants. They also improved relative water content (71.66%), gas exchange attributes, increased nutrients uptake, and osmolytes production of wheat. Up-regulation of antioxidant enzymes; superoxide dismutase (60.49%), peroxidase (55.99%), and catalase (81.69%) was also observed. This research work suggested that the application of SiO NPs and PGPR strains induced drought tolerance in wheat by modulating different physiological and metabolic processes in plants which ultimately improved the growth and yield of wheat under drought stress.Copyright © 2021 Elsevier Masson SAS. All rights reserved.
Waterlogging of winter crops at early and late stages: impacts on leaf physiology, growth and yield
DOI:10.3389/fpls.2018.01863
PMID:30619425
[本文引用: 1]
Waterlogging is expected to increase as a consequence of global climate change, constraining crop production in various parts of the world. This study assessed tolerance to 14-days of early-or late-stage waterlogging of the major winter crops wheat, barley, rapeseed and field pea. Aerenchyma formation in adventitious roots, leaf physiological parameters (net photosynthesis, stomatal and mesophyll conductances, chlorophyll fluorescence), shoot and root growth during and after waterlogging, and seed production were evaluated. Wheat produced adventitious roots with 20-22% of aerenchyma, photosynthesis was maintained during waterlogging, and seed production was 86 and 71% of controls for early-and late-waterlogging events. In barley and rapeseed, plants were less affected by early-than by late-waterlogging. Barley adventitious roots contained 19% aerenchyma, whereas rapeseed did not form aerenchyma. In barley, photosynthesis was reduced during early-waterlogging mainly by stomatal limitations, and by non-stomatal constraints (lower mesophyll conductance and damage to photosynthetic apparatus as revealed by chlorophyll fluorescence) during late-waterlogging. In rapeseed, photosynthesis was mostly reduced by non-stomatal limitations during early-and late-waterlogging, which also impacted shoot and root growth. Early-waterlogged plants of both barley and rapeseed were able to recover in growth upon drainage, and seed production reached ca. 79-85% of the controls, while late-waterlogged plants only attained 26-32% in seed production. Field pea showed no ability to develop root aerenchyma when waterlogged, and its photosynthesis (and stomatal and mesophyll conductances) was rapidly decreased by the stress. Consequently, waterlogging drastically reduced field pea seed production to 6% of controls both at early-and late-stages with plants being unable to resume growth upon drainage. In conclusion, wheat generates a set of adaptive responses to withstand 14 days of waterlogging, barley and rapeseed can still produce significant yield if transiently waterlogged during early plant stages but are more adversely impacted at the late stage, and field pea is not suitable for areas prone to waterlogging events of 14 days at either growth stage.
Vulnerability of photosynthesis and photosystem I in Jerusalem artichoke (Helianthus tuberosus L.) exposed to waterlogging
DOI:10.1016/j.plaphy.2018.02.017 URL [本文引用: 1]
Waterlogging stress in plants: unraveling the mechanisms and impacts on growth, development, and productivity
DOI:10.1016/j.envexpbot.2024.105824 URL [本文引用: 1]
Effect of waterlogging stress on grain nutritional quality and pod yield of peanut (Arachis hypogaea L.)
DOI:10.1111/jac.v209.2 URL [本文引用: 1]
Effects of high water table and short-term flooding on growth, yield, and seed quality of sunflower
DOI:10.1626/pps.14.233 URL [本文引用: 1]
A comparison of screening methods to identify waterlogging tolerance in the field in Brassica napus L. during plant ontogeny
DOI:10.1371/journal.pone.0089731 URL [本文引用: 1]
High sulfur (S) supplementation imparts waterlogging tolerance to oilseed rape (Brassica napus L.) through upregulating S metabolism and antioxidant pathways
DOI:10.1007/s00344-023-11034-8
[本文引用: 1]
Waterlogging significantly impacts plant growth and development by altering nutrient uptake and antioxidant enzyme functions, resulting in reduced yield. Plants need sulfur (S) to produce reduced glutathione (GSH), a thiol compound that combats abiotic stresses. It is hypothesized that supplying S to high S-demanding oilseed rape plants beyond its growth requirements can reduce the adverse effect of waterlogging stress. Therefore, this study evaluated the comparative effect of different S doses (mg kg− 1 soil), i.e., no-S (S0), low-S (S1, 35), medium-S (S2, 70), and high-S (S3, 140) on growth, yield and antioxidant defense systems of normal growing and waterlogged oilseed rape plants. Waterlogging was imposed at the inflorescence emergence stage for 7 days by retaining a 3-cm layer of water above the soil surface. Waterlogged plants supplemented with high-S showed improved growth and higher yield than those supplemented with lower S levels, and this response was associated with improved activity/contents of antioxidants, including ascorbate (AsA), GSH, ascorbate peroxidase, catalase, glutathione reductase, glutathione peroxidase, glutathione S-transferase, monodehydroascorbate reductase, and dehydroascorbate reductase, with concomitant lowering of hydrogen peroxide, dehydroascorbate and malondialdehyde content. Furthermore, leaf S concentration was enhanced in waterlogged plants treated with high-S, while anions were regulated whereby Cl− uptake was decreased. However, under control conditions, high-S did not provide additional benefits of improved plant growth, yield, and antioxidant activities compared to the medium-S application. Thus, it is concluded that additional S supplementation mitigates the adverse effects of waterlogging stress on oilseed rape, and needs to be employed as a potential strategy to alleviate the negative effects of this abiotic stress.
The roles of cell wall polysaccharides in response to waterlogging stress in Brassica napus L. root
DOI:10.1186/s12915-024-01972-4 [本文引用: 1]
Waterlogging tolerance of some tropical pasture grasses
Evaluating waterlogging stress response and recovery in barley (Hordeum vulgare L.): an image-based phenotyping approach
DOI:10.1186/s13007-024-01256-6 [本文引用: 2]
Data Processing system (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research
DOI:10.1111/ins.2013.20.issue-2 URL [本文引用: 1]
Comprehensive evaluation of high-oleic rapeseed (Brassica napus) based on quality, resistance, and yield traits: a new method for rapid identification of high-oleic acid rapeseed germplasm
DOI:10.1371/journal.pone.0272798
URL
[本文引用: 1]
To scientifically evaluate and utilize high-oleic acid rape germplasm resources and cultivate new varieties suitable for planting in the Hunan Province, 30 local high-oleic acid rape germplasms from Hunan were used as materials. The 12 personality indices of quality, yield, and resistance were comprehensively evaluated by variability, correlation, principal component, and cluster analyses. The results of variability showed that except for oleic acid, the lowest coefficient of variation was oil content, which was 0.06. Correlation analysis showed that oil content was positively correlated with main traits such as yield per plant and oleic acid, which could be used in the early screening of high-oleic rape germplasm. The results of principal component analysis showed that the 12 personality indicators were integrated into four principal components, and the cumulative contribution rate was 62.487%. The value of comprehensive coefficient ‘F’ was positively correlated with the first, second, and fourth principal components and negatively correlated with the third principal component. Cluster analysis showed that 30 high-oleic rape germplasms could be divided into four categories consisting of 9 (30%), 6 (20%), 7 (23%), and 8 (27%) high-oleic rape germplasms, each with the characteristics of \"high disease resistance\", \"high yield\", \"high protein\", and \"more stability\". This study not only provides a reference basis for high-oleic rape breeding but also provides a theoretical basis for their early screening.
Using cluster analysis and principal component analysis to group lines and determine important traits in white bean
DOI:10.1016/j.proenv.2015.07.145 URL [本文引用: 1]
Physiological mechanisms in response to waterlogging during seedling stage of Brassica napus L
DOI:10.3724/SP.J.1006.2024.34116 URL [本文引用: 1]
Conversion of lipids into carbohydrates rescues energy insufficiency in rapeseed germination under waterlogging stress
DOI:10.1111/ppl.v176.5 URL [本文引用: 1]
Physiological reaction differences of rapeseed (Brassica napus L.) with different waterlogging-tolerance to anoxia
Tissue-specific transcriptome and metabolome analysis reveals the response mechanism of Brassica napus to waterlogging stress
During the growth period of rapeseed, if there is continuous rainfall, it will easily lead to waterlogging stress, which will seriously affect the growth of rapeseed. Currently, the mechanisms of rapeseed resistance to waterlogging stress are largely unknown. In this study, the rapeseed (Brassica napus) inbred lines G230 and G218 were identified as waterlogging-tolerant rapeseed and waterlogging-sensitive rapeseed, respectively, through a potted waterlogging stress simulation and field waterlogging stress experiments. After six days of waterlogging stress at the seedling stage, the degree of leaf aging and root damage of the waterlogging-tolerant rapeseed G230 were lower than those of the waterlogging-sensitive rapeseed G218. A physiological analysis showed that waterlogging stress significantly increased the contents of malondialdehyde, soluble sugar, and hydrogen peroxide in rape leaves and roots. The transcriptomic and metabolomic analysis showed that the differential genes and the differential metabolites of waterlogging-tolerant rapeseed G230 were mainly enriched in the metabolic pathways, biosynthesis of secondary metabolites, flavonoid biosynthesis, and vitamin B6 metabolism. Compared to G218, the expression levels of some genes associated with flavonoid biosynthesis and vitamin B metabolism were higher in G230, such as CHI, DRF, LDOX, PDX1.1, and PDX2. Furthermore, some metabolites involved in flavonoid biosynthesis and vitamin B6 metabolism, such as naringenin and epiafzelechin, were significantly up-regulated in leaves of G230, while pyridoxine phosphate was only significantly down-regulated in roots and leaves of G218. Furthermore, foliar spraying of vitamin B6 can effectively improve the tolerance to waterlogging of G218 in the short term. These results indicate that flavonoid biosynthesis and vitamin B6 metabolism pathways play a key role in the waterlogging tolerance and hypoxia stress resistance of Brassica napus and provide new insights for improving the waterlogging tolerance and cultivating waterlogging-tolerant rapeseed varieties.
Waterlogging stress in cotton: damage, adaptability, alleviation strategies, and mechanisms
DOI:10.1016/j.cj.2020.08.005 URL [本文引用: 1]
Waterlogging causes early modification in the physiological performance, carotenoids, chlorophylls, proline, and soluble sugars of cucumber plants
DOI:10.3390/plants8060160
URL
[本文引用: 1]
Waterlogging occurs because of poor soil drainage and/or excessive rainfall and is a serious abiotic stress affecting plant growth because of declining oxygen supplied to submerged tissues. Although cucumber (Cucumis sativus L.) is sensitive to waterlogging, its ability to generate adventitious roots facilitates gas diffusion and increases plant survival when oxygen concentrations are low. To understand the physiological responses to waterlogging, a 10-day waterlogging experiment was conducted. The objective of this study was to measure the photosynthetic and key metabolites of cucumber plants under waterlogging conditions for 10 days. Plants were also harvested at the end of 10 days and analyzed for plant height (ht), leaf number and area, fresh mass (FM), dry mass (DM), chlorophyll (Chl), carotenoid (CAR), proline, and soluble sugars. Results indicated that cucumber plants subjected to the 10-day waterlogging stress conditions were stunted, had fewer leaves, and decreased leaf area, FM, and DM. There were differences in physiological performance, Chl, CAR, proline, and soluble sugars. Overall, waterlogging stress decreased net photosynthesis (A), having a negative effect on biomass accumulation. However, these decreases were also dependent on other factors, such as plant size, morphology, and water use efficiency (WUE) that played a role in the overall metabolism of the plant.
Short-term waterlogging has long-term effects on the growth and physiology of wheat
DOI:10.1046/j.0028-646X.2001.00318.x
URL
[本文引用: 1]
\n\n The effect of different durations of waterlogging and subsequent drainage is described for 3‐wk‐old wheat (Triticum aestivum) plants.
Morpho-anatomical and physiological responses to waterlogging of sesame (Sesamum indicum L.)
DOI:10.1016/j.plantsci.2013.03.014 URL [本文引用: 1]
Effects of waterlogging at different growth stages on physiological characteristics and seed yield of winter rape (Brassica napus L.)
DOI:10.1016/0378-4290(95)00075-5 URL [本文引用: 1]
Waterlogging events during stem elongation or flowering affect yield of oilseed rape (Brassica napus L.) but not seed quality
DOI:10.1111/jac.2018.204.issue-2 URL [本文引用: 1]
Effects of waterlogging at different growth stages on the stress-resistance physiological characteristics and yield formation of sweet potato
DOI:10.3724/SP.J.1006.2025.44116 URL [本文引用: 1]
Reactive oxygen species and antioxidant defense in plants under abiotic stress: revisiting the crucial role of a universal defense regulator
DOI:10.3390/antiox9080681
URL
[本文引用: 1]
Global climate change and associated adverse abiotic stress conditions, such as drought, salinity, heavy metals, waterlogging, extreme temperatures, oxygen deprivation, etc., greatly influence plant growth and development, ultimately affecting crop yield and quality, as well as agricultural sustainability in general. Plant cells produce oxygen radicals and their derivatives, so-called reactive oxygen species (ROS), during various processes associated with abiotic stress. Moreover, the generation of ROS is a fundamental process in higher plants and employs to transmit cellular signaling information in response to the changing environmental conditions. One of the most crucial consequences of abiotic stress is the disturbance of the equilibrium between the generation of ROS and antioxidant defense systems triggering the excessive accumulation of ROS and inducing oxidative stress in plants. Notably, the equilibrium between the detoxification and generation of ROS is maintained by both enzymatic and nonenzymatic antioxidant defense systems under harsh environmental stresses. Although this field of research has attracted massive interest, it largely remains unexplored, and our understanding of ROS signaling remains poorly understood. In this review, we have documented the recent advancement illustrating the harmful effects of ROS, antioxidant defense system involved in ROS detoxification under different abiotic stresses, and molecular cross-talk with other important signal molecules such as reactive nitrogen, sulfur, and carbonyl species. In addition, state-of-the-art molecular approaches of ROS-mediated improvement in plant antioxidant defense during the acclimation process against abiotic stresses have also been discussed.
The OsFBX388-OsFIP1-OsCatA module regulates ROS homeostasis and disease resistance in rice
DOI:10.1111/nph.70383
PMID:40650454
[本文引用: 1]
Plants are exposed to various biotic and abiotic environmental stresses, resulting in the generation of reactive oxygen species (ROS). One of the enzymes responsible for neutralizing excess ROS is catalase (CAT), which breaks down hydrogen peroxide (HO). However, the regulation of CAT in mediating HO homeostasis remains unclear. Here, we report that the DUF1644-containing protein OsFIP1 modulates HO homeostasis and positively regulates rice immunity. Overexpression (OE) of OsFIP1 leads to enhanced resistance against rice blast disease, accompanied by upregulation of defense-related genes, enhanced chitin-induced ROS burst and HO accumulation. Furthermore, OsFIP1 interacts with OsCatA. OE of OsFIP1 dramatically reduces accumulation of OsCatA in cytoplasm. Presence of OsFIP1 reduces OsCatA's tetramerization and CAT activity. Consistently, the knockout (KN) of OsCatA leads to enhanced resistance to rice blast. The E3 ligase OsFBX388 interacts with and ubiquitinates OsFIP1 for degradation. KN of OsFBX388 leads to overaccumulation of OsFIP1 and reduced CAT activity, thereby resulting in enhanced ROS burst and blast resistance. These findings highlight the crucial role of the OsFBX388-OsFIP1-OsCatA module in mediating ROS homeostasis and disease resistance, offering new insights into the mechanisms underlying immunity.© 2025 The Author(s). New Phytologist © 2025 New Phytologist Foundation.
Mechanisms of oxidative stress in plants: from classical chemistry to cell biology
DOI:10.1016/j.envexpbot.2014.06.021 URL [本文引用: 1]

{kind=link}
{kind=link}