


近年来,NRT2家族基因在多种植物中的功能研究已取得显著进展,在拟南芥中,AtNRT2.1和AtNRT2.2已被证实参与硝酸盐的高亲和性吸收[4],并在氮素缺乏条件下表现出显著的上调表达,AtNRT2.5在根部和成熟叶片中表达[5],在缺氮条件下能够调控硝酸盐的吸收和转运;在农作物中,NRT2家族基因的研究也逐渐深入,如小麦[6]中的TaNRT2.1和玉米[7]中的ZmNRT2.1均被证明对氮素吸收和利用效率有重要影响,ZmNRT2.1与NAR2蛋白相互作用后在硝酸盐的吸收途径中起主要作用[8],且主要在根部表达。然而,目前对StNRT2.5在马铃薯中的表达模式及具体功能研究较少,尤其是其在氮素吸收和利用中的作用机制尚不明确。
基于以上问题,本研究以30个不同马铃薯品种为材料,通过实时荧光定量PCR(qRT-PCR)分析了StNRT2.5在不同品种中的表达模式,并结合生物信息学分析对其保守结构域、系统发育关系及启动子元件进行了综合分析,旨在揭示StNRT2.5在不同马铃薯品种中的表达差异及其与氮素利用效率的关系,同时为深入理解StNRT2.5的功能及其在氮素吸收中的作用机制提供理论基础。此外,本研究也为马铃薯氮素利用效率的遗传改良提供了重要的参考依据,具有重要的理论和实践意义。
1 材料与方法
1.1 试验材料
30个马铃薯材料(表1)由吉林农业大学园艺学院马铃薯创新团队提供。选用MS基础培养基(主要氮源KNO3 1.9 g/L、NH4NO3 1.65 g/L),每瓶培养基扦插5株苗,每个处理10瓶。培养条件为光照强度6000 lx,光照时间14 h,温度25 ℃,湿度40%~60%的光照恒温培养箱中,21 d时采集30份马铃薯材料的叶(L)和根(R),重复3次,液氮冷冻后保存至-80 ℃冰箱。
表1 30个马铃薯品种信息
Table 1
| 编号Number | 品种Variety | 编号Number | 品种Variety |
|---|---|---|---|
| 1 | 龙薯14号 | 16 | 中薯6号 |
| 2 | 龙薯25号 | 17 | 中薯171 |
| 3 | 隆丰黑美人 | 18 | 中薯35号 |
| 4 | 克新30号 | 19 | 中薯2号 |
| 5 | 克新33号 | 20 | 中薯49号 |
| 6 | 克新28号 | 21 | 中薯48号 |
| 7 | 早大白 | 22 | 中薯45号 |
| 8 | 东农303 | 23 | 紫心 |
| 9 | 克新1号 | 24 | 春薯4号 |
| 10 | 春薯5号 | 25 | 春薯3号 |
| 11 | 富农 | 26 | 鄂薯3号 |
| 12 | 青薯24号 | 27 | 荷兰806 |
| 13 | AC142 | 28 | 延薯4号 |
| 14 | 荷兰14号 | 29 | 夏波蒂 |
| 15 | 大西洋 | 30 | 长崎 |
1.2 StNRT2.5表达模式分析
30份马铃薯材料分别提取根、叶RNA,反转录成cDNA后进行qRT-PCR。总RNA提取及cDNA合成:用TransZol Up Plus RNA Kit高纯度RNA提取试剂盒(ER501,北京全式金生物技术股份有限公司)提取马铃薯品种大西洋根和叶的总RNA,并用TransScript® All-in-One First-Strand cDNA Synthesis SuperMix for PCR反转录试剂盒(AU341,北京全式金生物技术股份有限公司)合成cDNA。
采用SnapGene软件设计qRT-PCR-StNRT2.5引物,内参基因引物EF1α由马铃薯创新团队提供,由吉林省生工生物有限公司合成。引物序列为:EF1α-F:GATGGTCAGACCCGTGAACA;EF1α-R:CCTTGGAGTACTTCG GGGTG[9]。荧光定量PCR按照Prime Script™ RT Master Mix(Perfect Real Time)试剂盒(北京全式金生物技术股份有限公司)说明书进行操作,反应体系(20 μL):TB Green Premix Ex Taq 10.0 μL,ROX Reference Dye 0.4 μL,cDNA 2.0 μL,上游引物0.8 μL,下游引物0.8 μL,ddH2O 6.0 μL。qRT-PCR反应程序:预变性95 °C 30 s;变性95 °C 5 s,退火60 °C 30 s,40个循环;95 °C 15 s,60 °C 1 min,95 ℃ 15 s。通过熔解曲线确认引物特异性,进行表达水平分析,将平均Ct值相对于内参归一化。采用2-∆∆Ct的方法进行分析,设置3次生物学重复。
1.3 StNRT2.5的克隆及生物信息学分析
基因筛选:参照课题组数据库中基因功能注释,筛选出StNRT2.5(LOC102586376),通过NCBI上进行BLAST比对,对序列进行分析,确定ORF框。RNA提取与cDNA合成:利用RNA提取试剂盒(ET111,北京全式金生物技术股份有限公司)提取马铃薯品种大西洋全株的总RNA,并使用反转录试剂盒(R312,北京全式金生物技术股份有限公司)合成cDNA第一链。PCR扩增:使用SnapGene 6.0.2设计特异性引物(StNRT2.5-F:ATGGAAA ACATGGATTTGGAATC;StNRT2.5-R:TATATTAG GAGTTCCATCTATAGGTG),进行目的片段PCR扩增,PCR产物回收纯化(EG101,北京全式金生物技术股份有限公司)。载体连接与转化:与TA/Blunt-Zero Cloning Kit载体(C601,南京诺唯赞生物科技股份有限公司)连接,将重组质粒转化到大肠杆菌感受态细胞DH5α(ZC101,北京庄盟国际生物基因科技有限公司)中,进行菌液PCR检测,具体操作方法参照说明书。鉴定正确的重组质粒送公司测序。生物信息学分析(表2)参考韩祉君等[10]的方法。
表2 生物信息学在线分析软件的名称及网址
Table 2
| 工具Tool | 网址URL | 功能Function |
|---|---|---|
| Blast | http://www.ncbi.nlm.nih.gov | 进行相似性比较 |
| ORF Finder | http://www.91bio.com/SMS2/filter_protein.html | 确定开放阅读框 |
| ProtParam | http://web.expasy.org/protparam/ | 计算蛋白理化性质 |
| ProScale | https://web.expasy.org/protscale/ | 预测亲水/疏水区 |
| Net Phos 3.1 Server | http://www.cbs.dtu.dk/services/NetPhos/ | 预测磷酸位点 |
| SignalP | http://www.cbs.dtu.dk/services/SignalP/ | 氨基酸信号肽预测 |
| TMHMM | http://www.cbs.dtu.dk/services/TMHMM/ | 氨基酸跨膜结构预测 |
| Interpro Scan | https://www.ebi.ac.uk/interpro/search/sequence/ | 蛋白质结构域预测 |
| Plant CARE | http://bioinformatics.psb.ugent/webtools/plantcare/html/ | 启动子顺式元件预测 |
| PSORT II Prediction | https://psort.hgc.jp/form2.html | 亚细胞定位预测 |
| String | https://string-db.org/ | 蛋白互作分析预测 |
| SOPMA | https://npsa-prabi.ibcp.fr/NPSA/npsa_sopma.html | 蛋白质二级结构预测 |
| Swiss-Model | https://swissmodel.expasy.org/interactive | 蛋白质三级结构预测 |
| Clustal X | http://www-igbmc.u-strasbg.fr/BioInfo/ | 多序列对比 |
| MEGA X | https://www.megasoftware.net/ | 构建系统进化树 |
2 结果与分析
2.1 StNRT2.5表达模式分析
根据StNRT2.5在不同马铃薯中的表达模式(图1),可以将30份马铃薯分为2类。一类是叶部表达量高于根部的品种,共有20份,包括隆丰黑美人、克新33号和长崎等,与延薯4号对氮素响应的转录分析一致[11];另一类是在根部表达高于叶部的品种,其中龙薯25号差异最显著。不同品种中StNRT2.5表达模式不同,使得在氮素吸收、转运和再分配过程中存在差异,影响其对氮素利用效率。在氮高效型马铃薯品种青薯和克新系列[12]中,StNRT2.5在叶片中维持较高表达,表明其在氮素的转运与再分配可能更为高效;而在氮低效型品种大西洋中,StNRT2.5在根部的表达量高于叶片,表明根系对氮素的代偿性吸收尝试,但氮素向地上部的转运与再分配受到限制,导致氮素无法被有效利用,从而表现为低氮利用效率。
图1
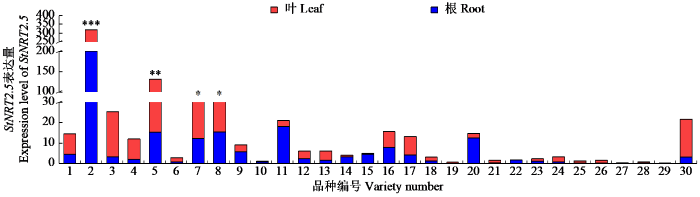
图1
StNRT2.5表达量分析
Fig.1
Expression level analysis of StNRT2.5
“*”: P < 0.05,“**”: P < 0.01,“***”: P < 0.001.
2.2 StNRT2.5的克隆及生物信息学分析
以马铃薯品种大西洋的cDNA为模板,利用特异性引物进行PCR扩增,得到大小约为1500 bp的序列(图2)。经测序,该片段大小为1512 bp。
图2
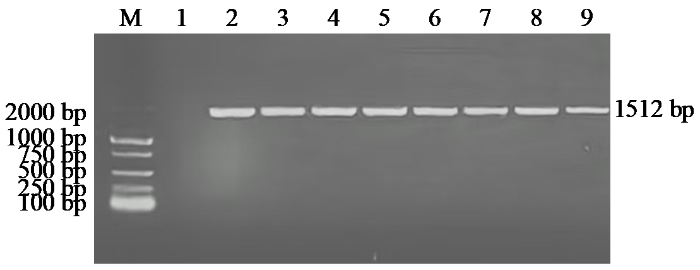
图2
StNRT2.5基因PCR扩增结果
M:DL2000;1:阴性对照;2:阳性对照;3~9:StNRT2.5菌液。
Fig.2
PCR amplification results of StNRT2.5 gene
M: DL2000; 1: negative control; 2: positive control; 3-9: StNRT2.5 bacterial solution.
表3 StNRT2.5蛋白理化性质分析
Table 3
| 蛋白 Protein | 氨基酸数量 Number of amino acids | 分子量 Molecular weight (Da) | 理论等电点 Theoretical isoelectric point | 原子数 Atomic number | 不稳定系数 Instability coefficient | 脂肪指数 Fat index | 亲水性平均值 Average hydrophilicity (GRAVY) | 负电荷残基 Negatively charged residues (Asp+Glu) | 正电荷残基 Positively charged residues (Arg+Lys) |
|---|---|---|---|---|---|---|---|---|---|
| StNRT2.5 | 503 | 54.73 | 8.68 | 7692 | 35.89 | 91.55 | 0.39 | 31 | 36 |
图3
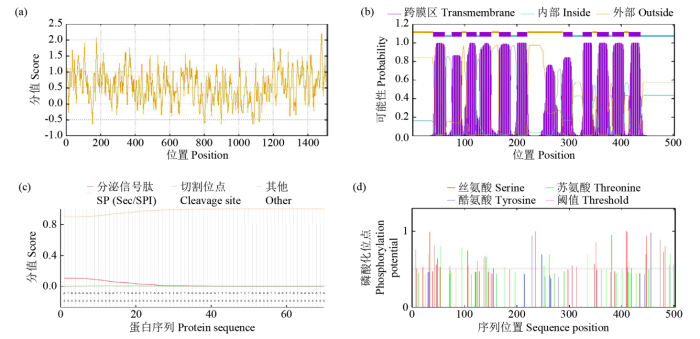
图3
StNRT2.5蛋白一级结构预测
(a) StNRT2.5蛋白亲疏水性分析;(b) StNRT2.5氨基酸跨膜结构预测;(c) StNRT2.5氨基酸信号肽;(d) StNRT2.5磷酸化位点预测。
Fig.3
Prediction of the primary structure of StNRT2.5 protein
(a) hydrophilicity hydrophobicity analysis of StNRT2.5 protein, (b) prediction of transmembrane structure of StNRT2.5 amino acids, (c) prediction of StNRT2.5 amino acid signal peptide, (d) prediction of StNRT2.5 phosphorylation site.
蛋白结构域搜索结果显示StNRT2.5蛋白同源超家族为MFS transporter(图4a),具有6个保守结构域,且均属于Nitrate transporter蛋白结构域,因此确定其为高亲和性硝酸盐转运蛋白。利用SOPMA预测StNRT2.5蛋白质二级结构,结果(图4b)显示StNRT2.5含有3种二级结构,其中α-螺旋占比最高。Swiss-Model三级结构分析(图4c)显示,StNRT2.5氨基酸序列与模板(AlphaFold DB model of A0A2Z7B1D0_9LAMI)覆盖率为80.24%,GMQE值为0.89。The BAR结果(图5)显示,eStNRT2.5在马铃薯根、叶与块茎中均有表达,且主要在叶片中表达。利用UniProt进行亚细胞定位预测,结果(图6)显示其定位在细胞膜中,因此推测StNRT2.5在细胞膜中行使其功能。
图4
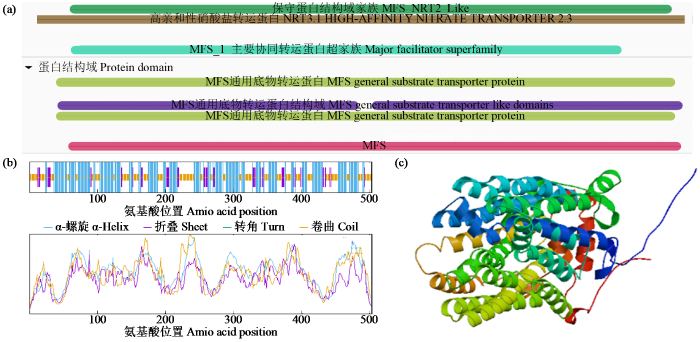
图4
StNRT2.5蛋白质结构域及二、三级结构分析
(a) StNRT2.5蛋白质结构域;(b) StNRT2.5的二级结构;(c) StNRT2.5的三级结构。
Fig.4
Analysis of domains as well as secondary and tertiary structure of StNRt2.5 protein
(a) StNRT2.5 protein domain, (b) secondary structure of StNRT2.5; (c) tertiary structure of StNRT2.5.
图5
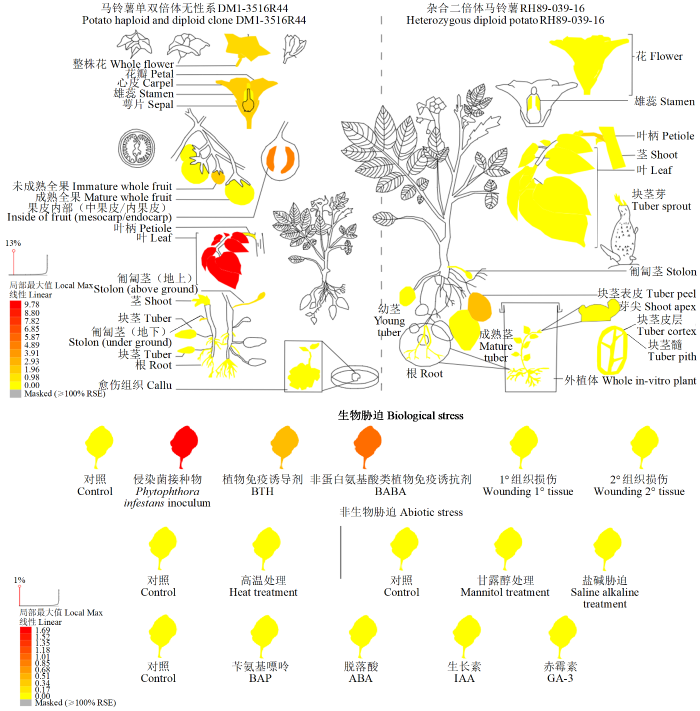
图6
对NRT2.5基因启动子区域(转录起始位点上游2000 bp)的顺式作用元件分析(图7)显示,该区域包含多种功能元件,涵盖光响应(如G-box、Box 4和LTR)、激素响应(如ABRE、ARE、CGTCA-motif和TCT-motif)、胁迫响应(如TC-rich repeats和CAAT-box)及基础转录调控(如TATA-box和CAAT-box)等类别,同时存在AE-box、A-box和AT-rich element等可能参与组织或发育特异性调控的元件。这些元件在启动子序列上的分布表明,NRT2.5基因的表达可能受光信号、多种植物激素及胁迫因素的综合调控,其启动子区域具有复杂的转录调控网络。氨基酸多重序列比对以拟南芥为参照,结果(图8)显示,利用NCBI-BlastX将StNRT2.5进行比对后使用MEGA软件构建StNRT2.5系统进化树,StNRT2.5与原始栽培的二倍体马铃薯(Solanum stenotomum)和多疣茄(Solanum verrucosum)中的StNRT2.5具有较高同源性,进化关系较为接近。
图7
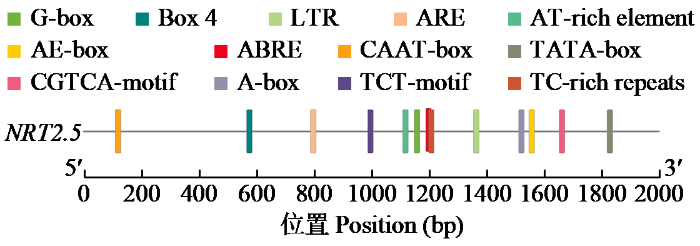
图8
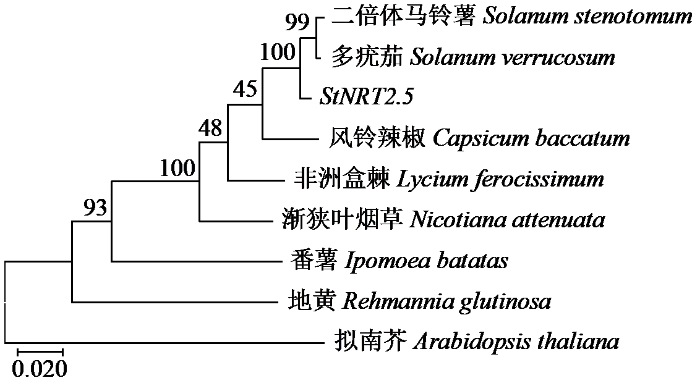
2.3 StNRT2.5互作蛋白研究
通过STRING数据库的分析,StNRT2.5蛋白被预测与多个已知的硝酸盐转运相关的基因存在互作关系。这些互作蛋白在氮代谢中发挥重要作用,可能与StNRT2.5的高亲和性转运功能形成协同作用。为了进一步验证这些互作关系,选择NIA2(硝酸还原酶,NR)和StNRT3.1作为目标互作蛋白,并与StNRT2.5进行蛋白互作预测(图9)。
图9
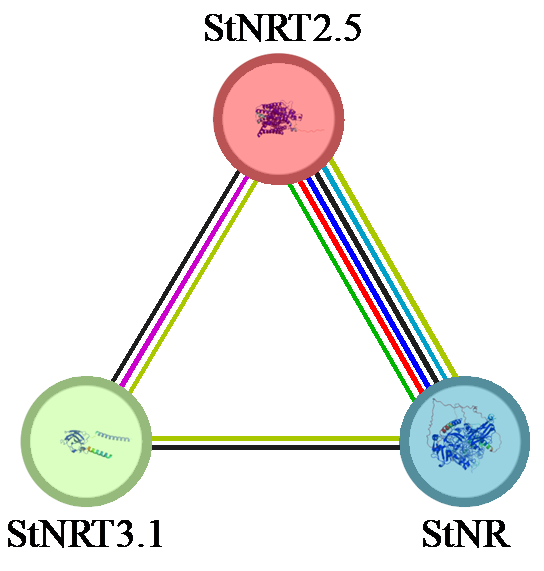
图9
StNRT2.5、StNR、StNRT3.1互作网络图
Fig.9
StNRT2.5, StNR, StNRT3.1 interaction network diagram
图10
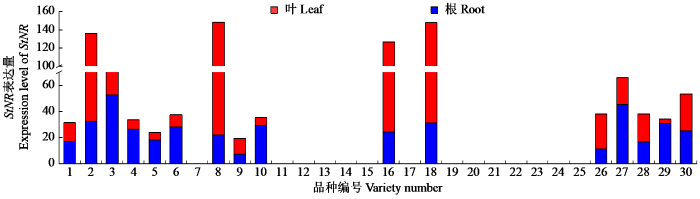
图11
3 讨论
高亲和性硝酸盐转运蛋白家族(NRT2)在氮匮乏条件下对植物氮素吸收和再分配中发挥着关键作用。30个马铃薯品种表达模式分析结果表明,StNRT2.5在不同品种中的表达存在组织差异性。主要可分为2类:一类是在叶片中高表达,推测其与AtNRT2.5在成熟叶片中表达一致,主要参与叶片中硝酸盐的再利用过程;另一类是在根部高表达,推测其与ZmNRT2.1与NAR2蛋白相互作用后表达一致,主要参与根部对硝酸盐的吸收过程。因此,经分析可知,StNRT2.5在根部的高表达有助于增加根部对氮素的吸收能力;而在叶片中高表达则有助于氮素从根部转运到地上部,在植株体内进行重新分配来满足不同组织器官对氮素的需求[15
生物信息学分析结果显示,StNRT2.5蛋白为稳定、疏水性蛋白。氨基酸序列比对结果表明,StNRT2.5基因编码的氨基酸序列含有MFS保守基序(MFS:G-XXX-D-XX-G-X-R)和硝酸盐/亚硝酸盐转运蛋白家族NNP保守基序(NNP:G-W/L-G- N-M/A-G),与硝酸盐转运蛋白NRT2基因家族的结构高度相似,说明StNRT2.5具有NRT2基因家族成员特征。启动子元件分析揭示了StNRT2.5启动子区域含有多种作用元件,这些元件的存在为StNRT2.5的研究提供了潜在的分子机制基础,并暗示该基因可能受到氮素信号和植物激素的协同调控。系统发育分析表明,StNRT2.5与原始栽培的二倍体马铃薯和多疣茄具有较高同源性,这一结果支持了StNRT2.5在进化上的保守性,表明其在不同茄科植物中可能具有相似的生物学功能。
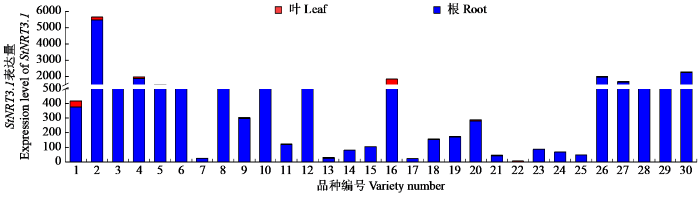
为了进一步验证StNRT2.5的功能,通过STRING数据库的分析,选择了StNR(NR)和StNRT3.1作为互作蛋白,并对其进行了蛋白互作预测及表达模式分析。结果表明StNR在叶片中表达更显著,其表达模式和受到基因型的显著影响,StNRT2.5与StNR的互作关系可能有助于将吸收的硝酸盐高效转运至叶片进行代谢,从而促进马铃薯植株的整体氮素利用效率[19];StNRT3.1在根系中高表达,与根系作为氮素吸收主要部位的功能相一致[20],且StNRT2.5与StNRT3.1的互作关系可能在硝酸盐的吸收过程中发挥关键作用,这种互作关系对于提高氮素吸收效率至关重要。本研究通过StNRT2.5的表达模式分析揭示了其在不同马铃薯品种和组织部位中的表达差异,并通过生物信息学分析揭示了StNRT2.5的保守结构域、进化关系以及启动子元件特征,为理解该基因在氮代谢过程中的调控机制研究提供更多参考,并利用STRING数据库分析预测了StNRT2.5与StNR、StNRT3.1的互作关系,为理解该基因在马铃薯氮素吸收中的功能提供了重要线索。这些结果不仅为后续的功能验证和分子机制研究奠定了基础,也为马铃薯氮素利用效率的遗传改良提供了理论支持。
4 结论
StNRT2.5的表达在不同品种中存在组织差异性,主要可分为2类:一类是在叶片中高表达,另一类则是在根部高表达;StNR在叶片中表达上调;StNRT3.1则主要在根系中表达。生物信息学分析表明StNRT2.5由503个氨基酸组成,为稳定、疏水性蛋白,具有典型的硝酸盐转运蛋白家族保守结构域,且StNRT2.5与原始栽培的二倍体马铃薯和多疣茄具有较高同源性。
参考文献
不同马铃薯品种的氮利用效率及其分类研究
DOI:10.3724/SP.J.1006.2019.84059
[本文引用: 1]

以7个马铃薯品种为供试材料, 设置大田条件下施氮和不施氮2种处理, 在对块茎产量和植株氮素吸收、利用评价的基础上, 将不同氮效率品种马铃薯分类并解析了其差异机制。基于2016年施氮和不施氮条件下各品种马铃薯的平均产量, 把不同氮效率品种马铃薯分为双高效型、低氮高效型、高氮高效型和双低效型。2017年选择双高效型、低氮高效型、双低效型的代表性品种, 对各类型氮效率差异进一步解析表明, 双高效型氮素利用效率显著高于另两个类型, 氮素吸收效率则是双高效型、低氮高效型显著高于双低效型。不施氮条件下, 双高效型马铃薯的干物质累积量在整个生育时期均显著高于另2个品种; 双高效型、低氮高效型氮素累积速率在出苗后0~50 d显著高于双低效型马铃薯。施氮条件下, 双高效型马铃薯的干物质累积量显著高于另2个品种, 与双低效型马铃薯相比, 双高效型和低氮高效型产量的提高主要归因于它们前期较高的干物质累积; 双高效型氮素累积速率显著高于双低效型、低氮高效型。双高效型马铃薯在各生育期的物质生产和氮素吸收能力强, 从而有利于氮效率提升和产量的形成。该研究结果可为马铃薯氮高效品种筛选和利用提供理论支撑。
硝酸盐转运蛋白NRT2在植物中的功能及分子机制研究进展
DOI:10.11983/CBB22134
[本文引用: 1]
氮素作为植物生长发育所需的大量元素, 对植物生长发育及作物产量具有重要作用。施入氮肥是植物及作物的主要氮素来源。面对当下过度施肥造成面源污染加剧的现状, 提高作物氮素利用效率, 实现“减肥增产”的绿色增产增效模式, 是促进我国农业可持续发展及保障国家粮食安全的重要措施。当土壤氮匮缺时, 硝酸盐转运蛋白NRT2家族成员对根系吸收及转运硝酸盐至关重要, 其中NRT2.1在植物缺氮时主要负责根部的硝酸根吸收。该文重点总结了模式植物拟南芥(Arabidopsis thaliana)及重要粮油作物中NRT2家族蛋白特别是NRT2.1的功能及调控机理研究进展, 旨在为后续挖掘NRT2在提高作物产量方面的潜力及分子调控机制研究提供重要依据。
Transgenic tobacco and tomato plants expressing Wasabi defensin genes driven by root- specific LjNRT2 and AtNRT2.1 promoters confer resistance against Fusarium oxysporum
A 150kDa plasma membrane complex of AtNRT2.5 and AtNAR2.1 is the major contributor to constitutive high‐affinity nitrate influx in Arabidopsis thaliana
DOI:10.1111/pce.2015.38.issue-8 URL [本文引用: 1]
Tobacco plants expressing the maize nitrate transporter ZmNrt2.1 exhibit altered responses of growth and gene expression to nitrate and calcium
DOI:10.1186/s40529-017-0203-9 URL [本文引用: 1]
Genome-wide identification and characterization of high-affinity nitrate transporter 2 (NRT2) gene family in tomato (Solanum lycopersicum) and their transcriptional responses to drought and salinity stresses
DOI:10.1016/j.jplph.2022.153684 URL [本文引用: 1]
Genome-wide identification,structural and gene expression analysis of the nitrate transporters (NRTs) family in potato (Solanum tuberosum L)
DOI:10.1371/journal.pone.0257383
URL
[本文引用: 1]
Nitrogen (N2) is the most important source of mineral N for plant growth, which was mainly transported by nitrate transporters (NRTs). However, little is known about the NRT gene family in potato. In this study, StNRT gene family members were identified in potato. In addition, we performed StNRT subfamily classification, gene structure and distribution analysis, and conserved domain prediction using various bioinformatics tools. Totally, 39 StNRT gene members were identified in potato genome, including 33, 4 and 2 member belong to NRT1, NRT2, and NRT3, respectively. These 39 StNRT genes were randomly distributed on all chromosomes. The collinearity results show that StNRT members in potato are closely related to Solanum lycopersicum and Solanum melongena. For the expression, different members of StNRT play different roles in leaves and roots. Especially under sufficient nitrogen conditions, different members have a clear distribution in different tissues. These results provide valuable information for identifying the members of the StNRT family in potato and could provide functional characterization of StNRT genes in further research.
Heterologous expression of nitrate assimilation related-protein DSNAR2.1/NRT3.1 affects uptake of nitrate and ammonium in nitrogen-starved Arabidopsis
DOI:10.3390/ijms21114027
URL
[本文引用: 1]
Nitrogen (N) is an essential macronutrient for plant growth. Plants absorb and utilize N mainly in the form of nitrate (NO3−) or ammonium (NH4+). In this study, the nitrate transporter DsNRT3.1 (also known as the nitrate assimilation-related protein DsNAR2.1) was characterized from Dianthus spiculifolius. A quantitative PCR (qPCR) analysis showed that the DsNRT3.1 expression was induced by NO3−. Under N-starvation conditions, the transformed Arabidopsis seedlings expressing DsNRT3.1 had longer roots and a greater fresh weight than the wild type. Subcellular localization showed that DsNRT3.1 was mainly localized to the plasma membrane in Arabidopsis root hair cells. Non-invasive micro-test (NMT) monitoring showed that the root hairs of N-starved transformed Arabidopsis seedlings had a stronger NO3− and NH4+ influx than the wild-type seedlings, using with NO3− or NH4+ as the sole N source; contrastingly, transformed seedlings only had a stronger NO3− influx when NO3− and NH4+ were present simultaneously. In addition, the qPCR analysis showed that the expression of AtNRT2 genes (AtNRT2.1–2.6), and particularly of AtNRT2.5, in the transformed Arabidopsis differed from that in the wild type. Overall, our results suggest that the heterologous expression of DsNRT3.1 affects seedlings’ growth by enhancing the NO3− and NH4+ uptake in N-starved Arabidopsis. This may be related to the differential expression of AtNRT2 genes.
玉米氮胁迫响应NRT基因的鉴定及ZmNRT2.5的克隆和表达分析
Genome-wide identification and functional analysis of nitrate transporter genes (NPF, NRT2 and NRT3) in maize
DOI:10.3390/ijms241612941
URL
[本文引用: 1]
Nitrate is the primary form of nitrogen uptake in plants, mainly transported by nitrate transporters (NRTs), including NPF (NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER FAMILY), NRT2 and NRT3. In this study, we identified a total of 78 NPF, seven NRT2, and two NRT3 genes in maize. Phylogenetic analysis divided the NPF family into eight subgroups (NPF1-NPF8), consistent with the results in Arabidopsis thaliana and rice. The NRT2 family appears to have evolved more conservatively than the NPF family, as NRT2 genes contain fewer introns. The promoters of all NRTs are rich in cis-acting elements responding to biotic and abiotic stresses. The expression of NRTs varies in different tissues and developmental stages, with some NRTs only expressed in specific tissues or developmental stages. RNA-seq analysis using Xu178 revealed differential expression of NRTs in response to nitrogen starvation and nitrate resupply. Moreover, the expression patterns of six key NRTs genes (NPF6.6, NPF6.8, NRT2.1, NRT2.5 and NRT3.1A/B) varied in response to alterations in nitrogen levels across distinct maize inbred lines with different nitrogen uptake rates. This work enhances our understanding of the structure and expression of NRTs genes, and their roles in nitrate response, paving the way for improving maize nitrogen efficiency through molecular breeding.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}