
作物杂志,2022, 第4期: 83–89 doi: 10.16035/j.issn.1001-7283.2022.04.012
甜荞茎秆重心高度和抗折力的遗传分析
胡丹( )
)
- 甘肃省种子总站,730030,甘肃兰州
Genetic Analysis of Culm Gravity Height and Snapping Resistance in Common Buckwheat
Hu Dan()
- Seed Administration Station of Gansu, Lanzhou 730030, Gansu, China
摘要:
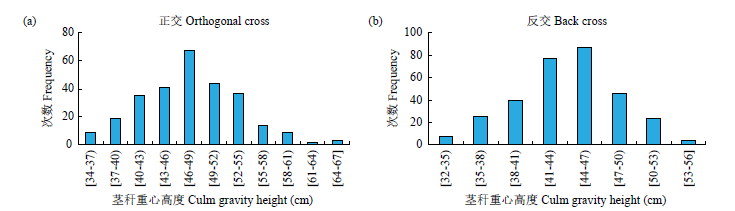
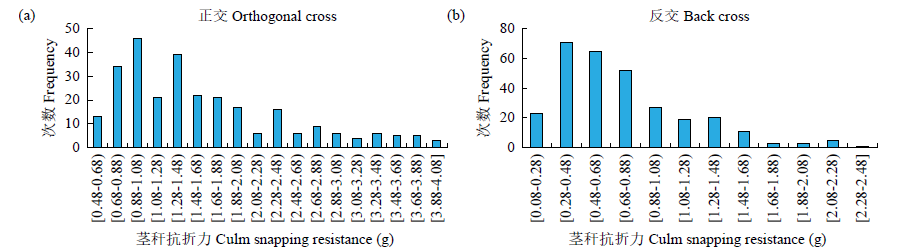
甜荞茎秆纤细和中空是造成甜荞极易倒伏的重要原因,茎秆重心高度和抗折力是影响甜荞倒伏的重要指标。以抗倒伏品种酉荞2号和易倒伏品种乌克兰大粒荞为亲本来配置正、反交组合。P1、P2、F1、B1、B2和F2群体茎秆重心高度和抗折力的遗传分析表明,茎秆重心高度最佳遗传模型为1对加性-显性主基因+加性-显性-上位性多基因模型和2对加性-显性-上位性主基因+加性-显性多基因模型,以加性效应为主,主基因遗传率大于多基因遗传率,环境变异大于遗传变异,可见环境对甜荞茎秆重心高度影响极大,可通过栽培措施降低甜荞茎秆重心高度,提高抗倒伏能力。茎秆抗折力最佳遗传模型为2对加性-显性-上位性主基因+加性-显性-上位性多基因模型和2对加性-显性-上位性主基因+加性-显性多基因模型,以加性效应为主,2对主基因间存在明显的基因互作效应,主基因遗传率大于多基因遗传率,在F2世代没有检测到多基因遗传率,主基因遗传率在F2世代最高,为88.94%,选择率高,可在早期世代进行选择来提高育种效率。


| [1] | 林汝法, 柴岩, 廖琴, 等. 中国小杂粮. 北京: 中国农业科技出版社, 2002. |
| [2] | 徐丽华, 潘宏, 赵英明. 荞麦——一种新兴的多用途作物. 国外农学-杂粮作物, 1998(3):51-53. |
| [3] | 张以忠, 陈庆富. 荞麦研究的现状与展望. 种子, 2004(3):39-42. |
| [4] | 张美莉. 萌发荞麦种子内黄酮与蛋白质的动态变化及抗氧化性研究. 北京: 中国农业大学, 2004. |
| [5] |
Awatsuhara R, Harada K, Maeda T. Antioxidative activity of the buckwheat polyphenol rutin in combination with ovalbumin. Molecular Medicine Reports, 2010, 3(1):121-125.
doi: 10.3892/mmr_00000228 pmid: 21472210 |
| [6] |
Griffith J Q, Couch J F, Lindauer M A. Effect of rutin on increased capillary fragility in man. Experimental Biology and Medicine, 1944, 55(3):228-229.
doi: 10.3181/00379727-55-14532 |
| [7] |
Jiang P, Burczynski F, Campbrll C, et al. Rutin and flavonoid contents in three buckwheat species Fagopyrum esculentum,F. tataricum, and F. homotropicum and their protective effects against lipid peroxidation. Food Research International, 2007, 40(3):356-364.
doi: 10.1016/j.foodres.2006.10.009 |
| [8] |
Wieslander G, Fabjan N, Vogrincic M, et al. Eating buckwheat cookies is associated with the reduction in serum levels of myeloperoxidase and cholesterol:a double blind crossover study in day-care centre staffs. Tohoku Journal of Experimental Medicine, 2011, 225(2):123-130.
pmid: 21931228 |
| [9] | Hagiwara M, Izusawa H, Inoue N, et al. Varietal differences of shoot growth characters related to lodging in Tartary buckwheat. Fagopyrum, 1999, 16:67-72. |
| [10] | 郭志利, 孙常青. 北方旱地荞麦抗倒栽培技术研究. 杂粮作物, 2007, 27(5):364-366. |
| [11] | 刘星贝, 汪灿, 易泽林, 等. 烯效唑干拌种对甜荞茎秆抗倒性能的影响. 作物学报, 2016, 42(1):93-103. |
| [12] | 刘星贝, 吴东倩, 易泽林, 等. 喷施烯效唑对甜荞茎秆抗倒性能及产量的影响. 中国农业科学, 2015, 48(24):4903-4915. |
| [13] | 汪灿, 阮仁武, 易泽林, 等. 荞麦茎秆解剖结构和木质素代谢及其与抗倒性的关系. 作物学报, 2014, 40(10):1846-1856. |
| [14] | 汪灿, 阮仁武, 易泽林, 等. 不同荞麦品种抗倒伏能力与根系及茎秆性状的关系. 西南大学学报(自然科学版), 2015, 37(1):65-71. |
| [15] |
Wang C, Hu D, Yi Z L, et al. Effects of uniconazole on the lignin metabolism and lodging resistance of culm in common buckwheat (Fagopyrum esculentum M.). Field Crops Research, 2015, 180:46-53.
doi: 10.1016/j.fcr.2015.05.009 |
| [16] | 佘恒志, 聂蛟, 易泽林, 等. 施硅量对甜荞倒伏及产量的影响. 中国农业科学, 2018, 51(14):2664-2674. |
| [17] | 邵庆勤. 叶面喷施多效唑提高小麦抗倒伏能力的生理机理研究. 南京:南京农业大学, 2017. |
| [18] | 杨文飞, 文廷刚, 孙爱侠, 等. 新型增产抗倒营养剂“劲丰谷德”对小麦抗倒性和产量的影响. 金陵科技学院学报, 2019, 35(1):65-68. |
| [19] | 袁圆, 汪波, 周广生, 等. 播期和种植密度对油菜产量和茎秆抗倒性的影响. 中国农业科学, 2021, 54(8):1613-1626. |
| [20] | 盖钧镒, 章元明, 王建康. 植物数量性状遗传体系. 北京: 科学出版社, 2003. |
| [21] | Stojsin R, 边静, 张满珍. 玉米籽粒产量和抗倒伏的重要茎秆性状的遗传. 杂粮作物, 1992(3):6-9. |
| [22] | 李得孝, 员海燕, 武玉华, 等. 玉米抗倒伏性状的遗传分析. 西北农业学报, 2004, 13(2):43-46. |
| [23] | 梁康迳, 林文雄. 水稻茎杆抗倒性的遗传及基因型×环境互作效应研究. 福建农业学报, 2000, 15(3):9-15. |
| [24] | 顾慧, 戚存扣. 甘蓝型油菜(Brassica napus L.)抗倒伏性状的主基因+多基因遗传分析. 作物学报, 2008, 34(3):376-381. |
| [25] | 李洪戈, 张丽萍, 伍晓明. 甘蓝型油菜茎秆强度性状的主基因+多基因遗传分析. 中国油料作物学报, 2018, 40(1):10-17. |
| [26] | 李英双, 胡丹, 易泽林, 等. 甜荞株高和茎粗的遗传分析. 作物学报, 2018, 44(8):1185-1195. |
| [27] | 张倩. 甘蓝型油菜主要株型性状的遗传分析和QTL初步定位. 重庆:西南大学, 2013. |
| [1] | 荣克伟, 柳波娟, 卢跃磊, 陈勇, 罗平, 赵康, 郝转芳, 高文伟. MAGIC群体的遗传特征及其在作物耐逆研究上的应用[J]. 作物杂志, 2022, (3): 9–19 |
| [2] | 凌晨, 刘洪, 杨哲, 黄展权, 陈孟强, 饶得花, 徐振江. 双季稻栽培对水稻DUS测试标准品种数量性状表达的影响[J]. 作物杂志, 2021, (4): 18–25 |
| [3] | 李忠南, 王越人, 张艳辉, 邬生辉, 曲海涛, 许正学, 李光发. 玉米DH系15D969超多穗行数的遗传分析[J]. 作物杂志, 2020, (5): 88–92 |
| [4] | 张晓玉,张亚玲,靳学慧,闫天雨,赵泽. 稻瘟病菌杂交后代致病性遗传分析[J]. 作物杂志, 2020, (2): 182–187 |
| [5] | 史娜溶,李静静,吴慧玉,孙道杰,冯毅,王辉,刘新伦,张玲丽. 西农979中长穗偃麦草(Thinopyrum ponticum)的遗传成分分析[J]. 作物杂志, 2019, (1): 15–21 |
| [6] | 张莉,李赞堂,王士银,麻艳超,东方阳,李学勇,徐江. 水稻氮素吸收低效型突变体osnad1的生理和遗传分析[J]. 作物杂志, 2018, (3): 68–76 |
| [7] | 张晓娟,周福平,张一中,邵强,范昕琦,刘勇,柳青山. 复粒高粱不育系的研究[J]. 作物杂志, 2017, (2): 72–75 |
| [8] | 杨修仕,郭忠贤,郭慧敏,王慧,刘三才. 播期和播量对荞麦产量及主要品质的影响[J]. 作物杂志, 2017, (1): 88–93 |
| [9] | 李忠南, 王克伟, 王越人, 邬生辉, 李光发. 玉米品种先玉335苗期叶绿素SPAD值的遗传分析[J]. 作物杂志, 2016, (4): 101–104 |
| [10] | 张海平,吴书峰,王志. 大豆对胞囊线虫4号生理小种的抗性遗传分析[J]. 作物杂志, 2016, (3): 21–26 |
| [11] | 刘杰,盛晋华,张雄杰. 甜荞产量与农艺性状和生理生化性状的灰色关联度分析[J]. 作物杂志, 2016, (1): 46–50 |
| [12] | 陈庆富, 陈其饺, 石桃雄, 等. 苦荞厚果壳性状的遗传及其与产量因素的相关性研究[J]. 作物杂志, 2015, (2): 27–31 |
| [13] | 刘化龙, 张宇, 邹德堂, 等. 香稻种质资源筛选及香味基因遗传研究[J]. 作物杂志, 2014, (6): 21–26 |
| [14] | 王欣欣, 卜一, 李尽朝, 等. 播种期对3个甜荞品种产量及主要性状的影响[J]. 作物杂志, 2014, (2): 110–113 |
| [15] | 侯迷红, 范富, 宋桂云, 等. 氮肥用量对甜荞麦产量和氮素利用率的影响[J]. 作物杂志, 2013, (1): 102–105 |
|
||