


薰衣草属唇形科薰衣草属(Lavandula),多年生半灌木或小灌木[1],是我国新疆特色的芳香植物,其植株耐旱、耐寒、喜光、怕涝,适宜质地疏松的土壤[2]。芳樟醇含量是评价薰衣草精油品质的重要指标之一[3]。芳樟醇合酶(linalool synthase,LIS)是由芳樟醇合酶基因编码而来,可以将异戊烯焦磷酸(IPP)和二甲基丙烯基二磷酸(DMAPP)的产物牻牛儿基焦磷酸(GPP)一步催化成芳樟醇,芳樟醇被乙醇酰基转移酶催化后可结合酰基辅酶A中的酰基形成乙酸芳樟酯[4]。天然的芳樟醇气味清甜、圆润,广泛存在于植物的挥发油中,在香精、香料和医疗保健产业上应用广泛[5]。Kessler等[6]证实芳樟醇活性挥发物具有杀虫的功效;刘静等[7]发现薰衣草精油具有明显的镇静催眠作用;Linck等[8]证实芳樟醇对小鼠也存在镇静作用。芳樟醇是薰衣草精油的重要功效组分,因此对芳樟醇在薰衣草体内合成机制的研究具有重要意义。
LIS基因最早从仙女扇中克隆,是第1个分离得到的单萜类合成酶基因[9]。国内外已经从烟草、仙女扇和莽山野柑等植物中克隆了LIS基因,研究[10]表明,其编码的蛋白质可催化GPP转化成芳樟醇。Mendoza-Poudereux等[11]将仙女扇LIS基因转入宽叶薰衣草后,转基因植株叶片中的芳樟醇含量显著增加,其含量是对照植株的10倍,且表型未发生变异。Lewinsohn等[12]将仙女扇LIS基因转入番茄中,转基因番茄中芳樟醇及其衍生物的含量也都有所上升。沉默烟草NaLIS基因,植株中芳樟醇含量显著下降[13]。将百合LiTPS2基因转入烟草植株中,转基因植株花中芳樟醇含量比野生型植株高2~3倍[14]。桂花OfTPS1和OfTPS2基因在烟叶中瞬时表达和稳定表达均可产生芳樟醇[15]。张雪荣[16]将从羽叶薰衣草中克隆的LIS基因转入矮牵牛中,检测到芳樟醇合酶。赵忠鑫等[17]对阔叶薰衣草中克隆的LIS基因进行了基因结构分析。
提高薰衣草精油中的芳樟醇含量是提升薰衣草精油品质的有效途径,研究薰衣草芳樟醇合酶的作用机理,可为分子水平上调控芳樟醇产量提供参考。本研究以高油杂薰衣草品种“杂花”(Lavandula x intermedia“Zahua”)为材料,克隆芳樟醇合酶基因LIS1和LIS2;以低油狭叶薰衣草品种“法国蓝”(Lavandula angustifolia“French blue”)为对照。比较了LIS1和LIS2基因在2个薰衣草品种不同发育时期的花冠和盛开期花器官不同组织的表达量,分析了2个基因原核表达和酶活性情况,初步揭示了它们在薰衣草萜类代谢通路上的调控作用,为利用基因工程手段培育精油高产、优质的薰衣草新品种提供了候选基因。
1 材料与方法
1.1 植物材料
供试薰衣草品种为“杂花”和“法国蓝”,于2018年6月在新疆伊犁地区70团采样。
1.2 载体和试剂
pET-28a(+)载体保存于新疆农业大学农业生物技术重点实验室;大肠杆菌感受态、荧光定量SuperMix、高保真酶、pEASY-T5载体和蛋白质Marker购自北京全式金生物技术有限公司;RNA提取试剂盒、胶回收试剂盒和质粒提取试剂盒购自北京天根生化科技有限公司;核酸染料和DNA Marker购自北京酷来搏科技有限公司;反转录试剂盒、限制性内切酶和T4 DNA连接酶购自赛默飞世尔科技有限公司;Ni-NTA蛋白纯化试剂盒、Elution Buffer、Binding/Wash Buffer和透析装置购自上海生工生物工程有限公司;GPP购自Sigma-Aldrich(上海)贸易有限公司。由昆泰锐生物技术有限责任公司进行引物合成和测序。LB培养基:每升培养基含胰蛋白胨10g、酵母提取物5g、NaCl 10g,固体LB培养基另加琼脂15g。HEPES缓冲液:称取0.598g HEPES、0.203g MgCl2、0.0016g MnCl2、0.00659g Na2WO4、0.00042g NaF和0.0077g DTT,加入10mL甘油,定容至100mL。
1.3 基因的克隆
根据NCBI提供的狭叶薰衣草(Lavandula angustifolia)芳樟醇合酶核苷酸序列(登录号:DQ263741.1)和阔叶薰衣草(Lavandula latifolia)芳樟醇合酶核苷酸序列(登录号:DQ421801.1)设计特异性引物F1、R1和F2、R2,引物序列见表1。提取“杂花”叶片RNA,反转录为cDNA作为模板,扩增目的基因。反应体系:cDNA 1μL、上下游引物各1μL、2.5mmol/L dNTP 5μL、10×EasyPfu Buffer 5μL、EasyPfu DNA Polymerase 1μL和H2O 36μL。引物F1和R1反应条件为94℃ 5min;94℃ 30s,60℃ 40s,72℃ 2min,35个循环;72℃ 10min;引物F2和R2反应条件为94℃ 5min;94℃ 30s,63℃ 40s,72℃ 2min,35个循环;72℃ 10min。PCR产物经1.5%琼脂糖凝胶电泳回收后,用pEASY-T5载体连接,菌液PCR鉴定阳性克隆,由昆泰锐生物技术有限责任公司测序。
表1 试验所用引物序列及用途
Table 1
| 引物 Primer | 序列 (5'-3') Sequence(5'-3') | 用途 Application |
|---|---|---|
| F1 | ATGTCTATCATTAGCATGCATGTGGG | 基因克隆 |
| R1 | TTAGGGATATGGCTCGAACATCAGGGT | 基因克隆 |
| F2 | ATGTCGATCAATATCAACATGCCTGC | 基因克隆 |
| R2 | TCATGCGTACGGCTCGAACA | 基因克隆 |
| F3 | AATAGGCCTGCAGCTTATAACCAT | 荧光定量 |
| R3 | TGGCGCGGCTTTGAAG | 荧光定量 |
| F4 | AATCGGCCGACTTCCTGAAT | 荧光定量 |
| R4 | TCGGAAACAAAGTTGTGGATTG | 荧光定量 |
| β-Actin-F | TGTGGATTGCCAAGGCAGAGT | 荧光定量内参 |
| β-Actin-R | AATGAGCAGGCAGCAACAGCA | 荧光定量内参 |
| F5 | CGGAATTCATGTCTATCATTAGCATGCA TGTGGG | 载体构建 |
| R5 | CCAAGCTTGGGATATGGCTCGAACATCA GGGT | 载体构建 |
| F6 | GGAATTCATATGATGTCGATCAATATCA ACATGCCTGC | 载体构建 |
| R6 | CGGAATTCTGCGTACGGCTCGAACA | 载体构建 |
1.4 生物信息学分析
使用表2中生物信息学网站对蛋白进行预测。
表2 生物信息学分析工具
Table 2
| 网站 Website | 网址 URL | 预测内容 Predict the content |
|---|---|---|
| ProtParam | 蛋白理化参数 | |
| InterProScan & Pfam | 蛋白功能结构域 | |
| DNAMAN | – | 氨基酸序列比对 |
| PSIPRED | 蛋白二级结构 | |
| SWISS-Model | 蛋白三级结构 | |
| MEGA 5.0 | – | 同源树构建 |
1.5 表达模式分析
“杂花”和“法国蓝”花器官不同发育时期的花冠各取3株,包括花蕾期、初开期、半开期、盛开期、衰败期以及花器官盛开期的不同组织(花萼、花冠、苞叶、雄蕊、雌蕊)(图1),提取RNA,反转录为cDNA,生物学重复3次。设计用于荧光定量表达分析的特异性引物F3、R3和F4、R4,引物序列见表1;以β-Actin(KY596032.1)基因为内参,利用荧光定量PCR技术分析LIS1和LIS2基因在薰衣草花器官不同发育时期花冠和盛开期不同组织中的表达模式,并比较基因在不同品种中的表达量。不同组织基因的表达量以叶片作为对照,不同发育时期的基因表达量以花蕾期作为对照,使用DPS软件处理数据,GraphPad Prism 5软件绘图。
图1

图1
薰衣草花器官不同组织和不同发育时期示意图
Fig.1
Schematic diagram of different tissues of lavender flower organs and different development stages
1.6 原核表达分析
1.6.1 重组蛋白表达条件优化 设计含有EcoR Ⅰ、Hind Ⅲ酶切位点的克隆引物F5和R5,以及含有Nde Ⅰ、EcoR Ⅰ酶切位点的克隆引物F6和R6,引物序列见表1,酶切扩增产物及pET-28a(+)载体,琼脂糖凝胶电泳后回收纯化目的基因及线性载体,用T4连接酶连接目的基因及线性载体后转化DH5α,挑选并鉴定阳性克隆,由昆泰锐生物技术有限责任公司测序,提取质粒转化Transetta(DE3)感受态。培养条件为37℃,220转/min,活化50μL pET-28a(+)-LIS1、pET-28a(+)-LIS2 Transetta (DE3)菌株至5mL LB培养基中,振荡培养12h,将活化后的500μL菌液加入15mL LB培养基中培养至OD600在0.4~0.6之间,分装到不同离心管中,每管分装1mL。每毫升菌液使用不同浓度IPTG(0.0、0.1、0.5、0.8、1.0、1.5和2.0mmol/L)的LB培养基,筛选不同诱导温度(28℃、37℃和40℃)和不同诱导时间(0、2、4、6和8h)进行条件优化。用60μL去离子水重悬菌液沉淀后加入15μL 5×蛋白上样缓冲液,100℃煮沸10min,进行SDS-PAGE凝胶电泳。
1.6.2 蛋白可溶性鉴定 采用已优化的诱导条件,活化菌株,扩大培养后取1mL作为诱导前全菌蛋白样品,其余菌液分为2组,一组加入诱导剂进行诱导,另一组不加诱导剂作为对照。将菌液8000转/min,4℃,离心10min,沉淀加入10mL PBS缓冲液,使用移液器吹打重悬后进行细胞破碎。破碎条件为200W,超声3s,间隔10s,柱头温度10℃。分别取60μL破碎后菌液作为未诱导全菌蛋白样品、诱导后全菌蛋白样品;取1mL诱导后破碎菌液,8000转/min,4℃,离心10min,取上清液60μL作为诱导后可溶样品,使用60μL去离子水重悬沉淀作为诱导后不可溶样品。分别取诱导前蛋白样品、未诱导蛋白样品、诱导后蛋白样品、诱导后可溶样品和诱导后不可溶样品于100℃煮沸10min,进行SDS-PAGE凝胶电泳。
1.6.3 溶解回收包涵体蛋白 大量制备包涵体蛋白后,参照上海生工生物工程有限公司Ni-NTA琼脂糖纯化树脂说明书,使用超声溶解包涵体后,取60μL包涵体溶解液,加入15μL 5×蛋白上样缓冲液,煮沸10min后备用。样品上柱后,收集流穿液,取流穿液60μL,加入15μL 5×蛋白上样缓冲液,煮沸10min后备用。用2mL洗脱缓冲液洗脱柱上的组氨酸标签蛋白2次,每次的洗脱液分别保存,取洗脱液60μL,加入15μL 5×蛋白上样缓冲液,100℃煮沸10min,进行SDS-PAGE凝胶电泳。随后采用透析法去除纯化蛋白中的咪唑。
1.7 酶活性检测
LIS1和LIS2重组蛋白酶活性鉴定反应体系为HEPES 500μL、GPP 20μL和重组蛋白200μL。向反应管中加入纯化后的蛋白,轻柔混匀,加入1mL正己烷封住液面,旋紧盖子,用封口膜密封后于30℃水浴30min,取出反应管。方法参照文献[18],略有改动。
将样品送至中国科学院新疆生态与地理研究所进行检测,采用气相色谱―质谱联用仪(GC-MS)分析样品成分。
2 结果与分析
2.1 LIS1和LIS2基因的克隆与序列分析
以“杂花”cDNA为模板,扩增LIS1和LIS2基因(图2)。使用DNAMAN软件进行分析,“杂花”LIS1基因的开放阅读框(ORF)长1809bp,编码一条由602个氨基酸组成的蛋白质序列;“杂花”LIS2基因的ORF长1695bp,编码一条由564个氨基酸组成的蛋白质序列。使用ProtParam在线程序计算“杂花”LIS1和LIS2基因编码蛋白的理化参M:Marker Ⅶ数,结果表明,“杂花”LIS1蛋白质等电点为5.45,分子量约为70.32kDa;“杂花”LIS2蛋白等电点为5.08,分子量约为65.65kDa。“杂花”LIS1和LIS2芳樟醇合酶基因序列相似度为58.54%。
图2
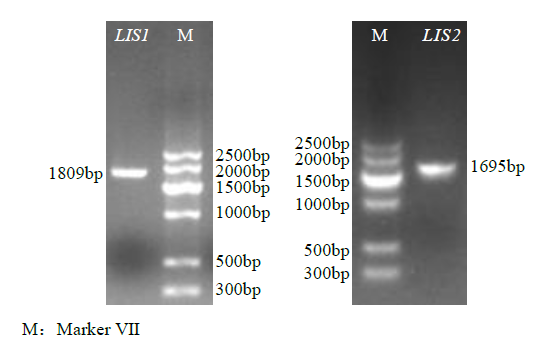
图2
LIS基因扩增产物琼脂糖凝胶电泳
Fig.2
Agarose gel electrophoresis of LIS gene amplification products
2.2 LIS1和LIS2蛋白质生物信息学分析
2.2.1 LIS1和LIS2蛋白功能结构域预测 使用EMBL的Pfam在线软件推导出LIS1和LIS2蛋白功能结构域(图3),可看出2种蛋白质序列含有Terpene_synth结构域和Terpene_synth_C结构域,预测LIS1和LIS2蛋白属于Terpene_synth超家族和Isoprenoid_Biosyn_C1超家族。
图3

2.2.2 LIS1和LIS2蛋白结构预测 使用PSIPRED在线工具预测LIS1和LIS2蛋白二级结构。LIS1蛋白二级结构包含3个β-折叠和28个α-螺旋;LIS2蛋白二级结构包含29个α-螺旋(图4)。
图4
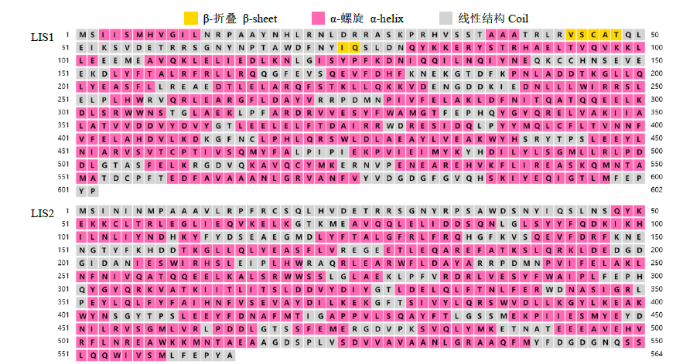
图4
LIS1和LIS2蛋白二级结构预测
Fig.4
Predicted secondary structure of LIS1 and LIS2 proteins
使用SWISS-Model在线程序的自动模式对LIS1和LIS2蛋白的三级结构进行预测分析,发现“杂花”LIS1和LIS2蛋白三级结构中分别含有GCC元件结合结构域SMTL ID 2o1x.1 B和SMTL ID 1n1b.1 B,相似性分别达到52.50%和64.56%(图5)。
图5

图5
LIS1和LIS2蛋白的三级结构预测
Fig.5
Predicted tertiary structures of LIS1 and LIS2 proteins
2.2.3 LIS1和LIS2蛋白进化分析 使用DNAMAN软件对水薄荷(Mentha aquatica)、甜舌草(Phyla dulcis)、陆地棉(Gossypium hirsutum)、紫苏(Perilla setoyensis)、一串红(Salvia splendens)、桂花(Osmanthus fragrans var. thunbergii)、阔叶薰衣草、狭叶薰衣草和杂薰衣草中的芳樟醇合酶蛋白进行相关蛋白多序列比对,发现LIS1和LIS2蛋白序列具有单萜类合成酶的所有保守氨基酸和功能高度保守域序列,即DDXXD、RRX8W和DDXXTXXXE功能基团(图6)。使用MEGA 5.0软件邻近连接法对上述物种的芳樟醇合酶蛋白进行相关蛋白的同源树构建,结果(图7)显示,LIS1蛋白和阔叶薰衣草、一串红中的芳樟醇合酶亲缘关系相近;LIS2蛋白和狭叶薰衣草、杂薰衣草中的芳樟醇合酶亲缘关系相近。
图6
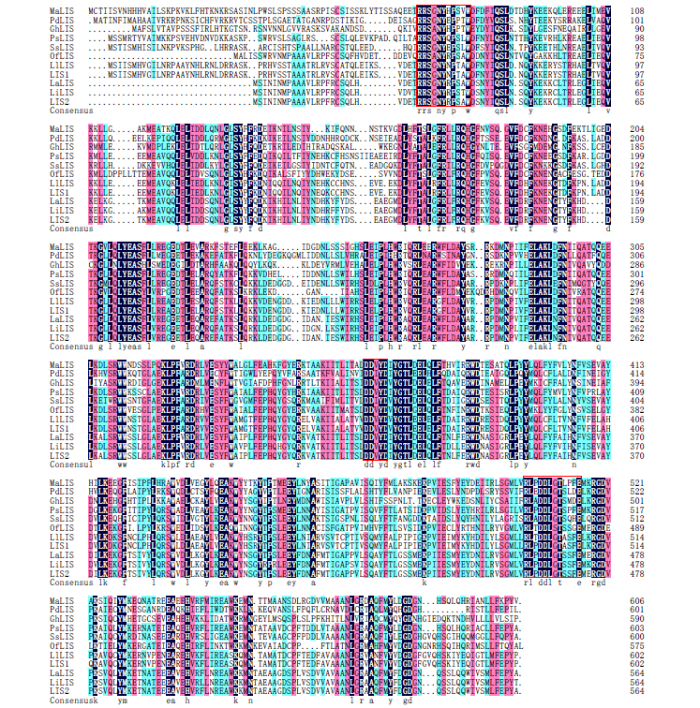
图6
LIS1和LIS2蛋白与其他物种芳樟醇合酶蛋白的多序列比对
第1个红色框线为RRX8W功能基团,第2个红色框线为DDXXD功能基团,第3个红色框线为DDXXTXXXE功能基团
Fig.6
Multiple alignment of amino acid sequences of LIS1 and LIS2 proteins with linalool synthase proteins from other species
The first red box line is RRX8W functional group, the second red box line is DDXXD functional group, and the third red box line is DDXXTXXXE functional group
图7

图7
LIS1和LIS2蛋白的系统进化树分析
分支点的数值表示bootstrap验证中基于1000次重复的置信度
Fig.7
Phylogenetic tree analysis of LIS1 and LIS2 proteins
The numerical values of the branch points represent the confidence of the bootstrap validation based on 1000 repetitions
2.3 LIS1和LIS2基因的表达模式分析
经实时荧光定量PCR分析,在2个薰衣草品种不同发育时期、花器官不同组织中均检测到了LIS1和LIS2基因的表达(图8-9)。在花器官不同发育时期,LIS1基因在“杂花”花冠的衰败期表达量最高,在花蕾期和初开期的表达量相对较低;该基因在“法国蓝”花冠中的初开期、半开期、盛开期和衰败期表达量缓慢上升。比较LIS1基因在2个薰衣草品种中不同时期表达量情况,该基因在花蕾期和初开期相对表达量差异不大,在半开期、盛开期和衰败期“杂花”的表达量均高于“法国蓝”;LIS2基因在“杂花”花冠的不同发育时期表达量逐渐上升,衰败期达到最高,但“法国蓝”在半开期表达量达到最高,之后盛开期、衰败期逐渐下降。
图8

图8
LIS1基因表达量及显著性分析
不同字母表示在0.05水平上差异显著,下同
Fig.8
LIS1 gene expression level and significance analysis
Different letters indicate significant difference at 0.05 level, the same below
图9

在花器官不同组织中,LIS1基因在“杂花”花萼中表达量最高,其次为花冠,在花蕊中的相对表达量较低,而该基因在“法国蓝”花器官各组织中几乎不表达;LIS1基因在“杂花”花器官不同组织中的相对表达量明显高于其在“法国蓝”花器官不同组织中的(雌蕊、雄蕊和叶片除外)相对表达量;LIS2基因在“杂花”花萼中表达量最高,其次为花冠,在其他组织中表达量较低;LIS2基因在“法国蓝”雄蕊中表达量最高,其次为花萼。LIS2基因在“法国蓝”雄蕊中的表达量明显高于其在“杂花”中的表达量,在花冠、花萼和苞叶中的表达量低于在“杂花”中的表达量。LIS1和LIS2基因在2个品种花器官中的表达量均存在差异,LIS2基因在2个品种花器官不同发育时期、不同组织中的相对表达量均高于LIS1基因在2个品种中的表达量。
2.4 LIS1和LIS2基因原核表达分析
构建pET-28a(+)-LIS1和pET-28a(+)-LIS2重组质粒,经双酶切后,采用琼脂糖凝胶电泳检测酶切产物,结果(图10)表明,酶切后获得的条带与目的条带大小一致。测序结果与目的序列一致,证明载体构建成功。
图10

图10
重组质粒pET-28a(+)-LIS1和pET-28a(+)-LIS2的双酶切鉴定
M:Marker 15 000;1:pET-28a(+)-LIS1质粒经EcoRⅠ、Hind Ⅲ双酶切;2:pET-28a(+)-LIS1线性质粒;3:pET-28a(+)-LIS2质粒经NdeⅠ、EcoRⅠ双酶切;4:pET-28a(+)-LIS2线性质粒
Fig.10
Identification of recombinant plasmid pET-28a(+)-LIS1 and pET-28a(+)-LIS2 by double enzyme digestion
M: Marker 15000; 1: pET-28a(+)- LIS1 digested by double enzyme EcoRⅠand Hind Ⅲ; 2: pET-28a(+)-LIS1 linear plasmid; 3: pET-28a(+)- LIS2 digested by double enzyme NdeⅠ and EcoRⅠ; 4: pET-28a(+)-LIS2 linear plasmid
活化Transetta/pET-28a(+)-LIS1和Transetta/pET-28a(+)-LIS2表达菌后,对诱导剂浓度、诱导时间和诱导温度进行优化,Transetta/pET-28a(+)-LIS1在37℃、IPTG终浓度1mmol/L时,诱导4h后,LIS1蛋白表达量最大,Transetta/pET-28a(+)-LIS2在37℃、IPTG终浓度1mmol/L时,诱导6h后,LIS2蛋白表达量最大。取细菌沉淀进行全菌蛋白上样,进行SDS-PAGE凝胶电泳,结果(图11)表明,诱导蛋白条带大小与预测蛋白一致。
图11

图11
LIS1和LIS2重组蛋白原核表达分析
M:蛋白ProteinRuler®Ⅰ,下同;1:pET-28a(+)未诱导;2:pET-28a(+)诱导后;3:pET-28a(+)-LIS2蛋白诱导前;4:pET-28a(+)-LIS2蛋白未诱导;5:pET-28a(+)-LIS2蛋白诱导后;6:pET-28a(+)未诱导;7:pET-28a(+)诱导后;8:pET-28a(+)-LIS1蛋白诱导前;9:pET-28a(+)-LIS1蛋白未诱导;10:pET-28a(+)-LIS1蛋白诱导后
Fig.11
Prokaryotic expression analysis of LIS1 and LIS2 recombinant proteins
M: ProteinRuler®Ⅰ, the same below; 1: transetta with pET-28a(+) uninduced; 2: transetta with pET-28a(+) induced by IPTG; 3: transetta with pET-28a(+)-LIS2 before induction; 4: transetta with pET-28a(+)-LIS2 uninduced; 5: transetta with pET-28a(+)-LIS2 by IPTG; 6: transetta with pET-28a(+) uninduced; 7: transetta with pET-28a(+) induced by IPTG; 8: transetta with pET-28a(+)-LIS1 before induction; 9: transetta with pET-28a(+)-LIS1 uninduced; 10: transetta with pET-28a(+)-LIS1 by IPTG
对重组蛋白LIS1和LIS2分别进行可溶性鉴定,发现二者均为不可溶的包涵体蛋白(图12)。
图12
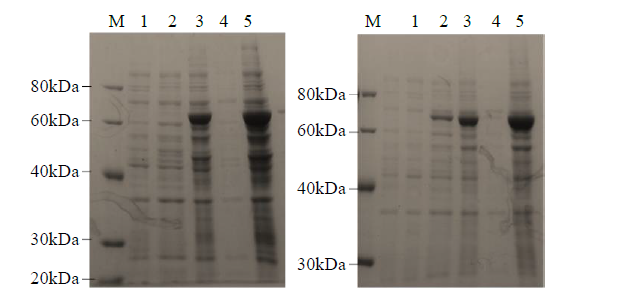
图12
LIS1和LIS2重组蛋白可溶性鉴定
1:诱导前蛋白样品;2:未诱导蛋白样品;3:诱导后蛋白样品;4:诱导后可溶样品;5:诱导后不可溶样品
Fig.12
Soluble identification of LIS1 and LIS2 recombinant proteins
1: protein samples before induction; 2: non-induced protein sample; 3: protein samples after induction; 4: soluble sample after induction; 5: insoluble sample after induction
对重组蛋白LIS1和LIS2分别进行溶解回收鉴定,结果(图13)表明,回收后的蛋白条带均可见,符合进行体外酶活性检测的条件。
图13

图13
LIS1、LIS2重组蛋白回收鉴定
1:包涵体溶解液;2:溶解蛋白流穿液;3:溶解蛋白洗脱液1;4:溶解蛋白洗脱液2
Fig.13
Recovery identification of LIS1 and LIS2 recombinant proteins
1: inclusion body dissolution solution; 2: dissolved protein fluid; 3: dissolved protein eluent 1; 4: dissolved protein eluent 2
图14
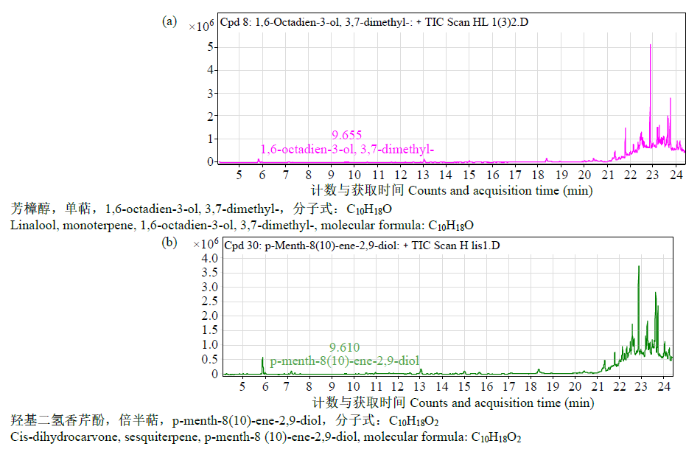
图14
LIS1重组蛋白添加底物输出产物GC-MS检测
Fig.14
GC-MS detection of output products of LIS1 recombinant protein supplemented with substrate
图15

图15
LIS2重组蛋白添加底物输出产物GC-MS检测
芳樟醇,单萜,1,6-octadien-3-ol, 3,7-dimethyl-,分子式:C10H18O
Fig.15
GC-MS detection of output products of LIS2 recombinant protein supplemented with substrate
Linalool, monoterpene, 1,6-octadien-3-ol, 3,7-dimethyl-, molecular formula: C10H18O
3 讨论
3.1 芳樟醇合酶基因的结构
芳樟醇合酶是植物中合成芳樟醇的主要催化酶,萜烯合成酶氨基酸序列的相似性并不意味着功能的相似性,芳樟醇合酶可以独立于其他萜类合成酶基因自由地进化和分离[19]。本研究克隆了“杂花”薰衣草的芳樟醇合酶基因LIS1和LIS2,在氨基酸序列中均发现了单萜类合成酶的所有保守氨基酸序列和功能高度保守域序列,包括结合二价金属离子的DDXXD基团、具有环化异构功能的RRX8W基团和结合固定底物的DDXXTXXXE基团。本研究中2条芳樟醇合酶序列相似度为58.54%,推测结构差异是造成LIS1和LIS2蛋白质催化底物GPP合成产物数量不一致的原因,该结果与Cseke等[20]认为LIS是一种复合基因,可能由2种不同类型的萜烯合成酶基因重组演变而来相一致。Raguso等[21]也验证了芳樟醇合酶与柠檬烯合成酶(LMS)和古巴酯焦磷酸合成酶(CPS)基因具有相同序列。
3.2 芳樟醇合酶基因表达模式及差异
在同一物种内,芳樟醇合酶基因在不同品种或生态型表达水平存在差异[22]。Lin等[23]发现,不同品种肉桂的芳樟醇合酶表达存在差异,芳樟醇型品种基因表达水平高于肉桂醛型品种和樟脑型品种。在“杂花”和“法国蓝”2个品种花器官不同发育时期和不同组织中,LIS2基因的表达量均高于LIS1,且LIS1基因在“杂花”中表达量偏高,LIS2基因在“法国蓝”中的表达量相对偏高。芳樟醇合酶基因在同一植株不同发育时期和不同组织的表达水平也存在差异。如姜花中芳樟醇合酶基因在营养器官中几乎不表达,但在花器官不同发育时期的表达水平与芳樟醇含量呈正相关[24]。小苍兰FhTPS1基因在花器官不同发育时期和不同组织中均表现出不同的表达模式[25]。铁线莲萼片中芳樟醇合酶基因的表达水平高于其在花瓣和子房中的表达水平[26]。在蓍草不同组织中LIS基因表达存在差异,该基因在叶和腺毛中的表达量较高,而在花器官中则不表达。本研究中,LIS1基因在“法国蓝”不同发育时期的表达量随时间呈现递增趋势,但在2个品种不同组织中该基因的表达量极低;LIS2基因在“杂花”不同发育时期的表达量随着时间呈现递增趋势,在2个品种的不同组织中该基因表达量较高。薰衣草花穗中的精油大部分包含在花萼的腺体中[28],LIS1基因在“杂花”花萼中表达量最高,但在“法国蓝”花萼中几乎不表达;LIS2基因在“杂花”和“法国蓝”花萼中的表达量均处于较高水平,推测其是合成花萼中精油成分的主要萜类合成酶;LIS2基因在“法国蓝”半开期、盛开期和衰败期表达量没有显著差异。为进一步揭示2个基因调控芳樟醇合成的作用,还需将基因表达模式与花器官不同发育时期和不同组织的芳樟醇含量相结合进行一致性分析。
3.3 芳樟醇合酶的原核表达和活性鉴定
本研究利用大肠杆菌表达重组蛋白,纯化回收蛋白后进行酶活性鉴定,结果表明,LIS1和LIS2蛋白都具有单萜合酶活性,LIS1蛋白参与合成芳樟醇和5-[1-羟甲基乙烯基]2-甲基环己醇,LIS2蛋白参与合成芳樟醇。研究[33]表明,部分植物单萜合酶在特异的代谢环境中会产生多种产物,LIS1蛋白在体外活性鉴定中催化合成了2种单萜类物质,推测其在植物体内可催化合成多种产物。重组蛋白的酶活鉴定结果初步揭示了2个基因具有催化底物合成芳樟醇的功能,但仍需通过转基因技术在植物体内进行过表达和基因沉默,以期准确验证其功能。
4 结论
对从“杂花”薰衣草中克隆的LIS1和LIS2基因进行了生物信息学分析。结果表明,薰衣草LIS1和LIS2基因是萜类合成酶中的成员。LIS1和LIS2基因在不同薰衣草品种和花器官不同组织、不同发育时期的表达量均存在差异。重组蛋白的体外酶活性鉴定表明,LIS1和LIS2基因编码的蛋白可催化底物GPP生成芳樟醇,同时LIS1蛋白还可催化底物GPP生成5-[1-羟甲基乙烯基]2-甲基环己醇。初步揭示了薰衣草LIS1和LIS2基因具有调控芳樟醇生物合成的功能。
参考文献
Biosynthesis and therapeutic properties of Lavandula essential oil constituents
DOI:10.1055/s-0030-1250136
PMID:20665367
[本文引用: 1]

Lavenders and their essential oils have been used in alternative medicine for several centuries. The volatile compounds that comprise lavender essential oils, including linalool and linalyl acetate, have demonstrative therapeutic properties, and the relative abundance of these metabolites is greatly influenced by the genetics and environment of the developing plants. With the rapid progress of molecular biology and the genomic sciences, our understanding of essential oil biosynthesis has greatly improved over the past few decades. At the same time, there is a recent surge of interest in the use of natural remedies, including lavender essential oils, in alternative medicine and aromatherapy. This article provides a review of recent developments related to the biosynthesis and medicinal properties of lavender essential oils.© Georg Thieme Verlag KG Stuttgart · New York.
Defensive function of herbivore-induced plant volatile emissions in nature
Herbivore attack is known to increase the emission of volatiles, which attract predators to herbivore-damaged plants in the laboratory and agricultural systems. We quantified volatile emissions from Nicotiana attenuata plants growing in natural populations during attack by three species of leaf-feeding herbivores and mimicked the release of five commonly emitted volatiles individually. Three compounds (cis-3-hexen-1-ol, linalool, and cis-alpha-bergamotene) increased egg predation rates by a generalist predator; linalool and the complete blend decreased lepidopteran oviposition rates. As a consequence, a plant could reduce the number of herbivores by more than 90% by releasing volatiles. These results confirm that indirect defenses can operate in nature.
Inhaled linalool-induced sedation in mice
DOI:10.1016/j.phymed.2008.08.001 URL [本文引用: 1]
Purification and characterization of S-linalool synthase,an enzyme involved in the production of floral scent in Clarkia breweri
S-Linalool is one of the volatiles emitted by Clarkia breweri Grey [Green] flowers to attract its moth pollinator. S-Linalool synthase, the enzyme that stereoselectively converts the ubiquitous C10 intermediate GPP to S-linalool, is abundant in stigmata of freshly opened flowers, and it was purified to > 95% homogeneity by anion-exchange and hydroxyapatite chromatography. S-Linalool synthase is operationally soluble as are other monoterpene synthases, has a Km of 0.9 microM for geranyl pyrophosphate, exhibits a strict requirement for a divalent metal cofactor with a preference for Mn2+ (Km = 45 microM), and shows an optimal pH of 7.4. The enzyme is active as a monomer of 76 +/- 3 kDa as determined by gel permeation chromatography and polyacrylamide gel electrophoresis. Neither S- nor R-linalyl pyrophosphates are substrates for the C. breweri S-linalool synthase, although this tertiary allylic pyrophosphate ester is a bound intermediate in the biosynthesis of cyclic monoterpenes from geranyl pyrophosphate in many plant species, where it also serves as an alternate substrate.
Evolution of floral scent in Clarkia:novel patterns of S-linalool synthase gene expression in the C. breweri flower
Flowers of Clarkia breweri, an annual plant from the coastal range of California, emit a strong sweet scent of which S-linalool, an acyclic monoterpene, is a major component. Chromosomal, chemical, and morphological data, and the species' geographic distribution, suggest that C. breweri evolved from an extant nonscented species, C. concinna. A cDNA of Lis, the gene encoding S-linalool synthase, was isolated from C. breweri. We show that in C. breweri, Lis is highly expressed in cells of the transmitting tract of the stigma and style and in the epidermal cells of petals, as well as in stamens, whereas in the nonscented C. concinna, Lis is expressed only in the stigma and at a relatively low level. In both species, changes in protein levels parallel changes in mRNA levels, and changes in enzyme activity levels parallel changes in protein levels. The results indicate that in C. breweri, the expression of Lis has been upregulated and its range enlarged to include cells not expressing this gene in C. concinna. These results show how scent can evolve in a relatively simple way without the evolution of highly specialized "scent glands" and other specialized structures. Lis encodes a protein that is structurally related to the family of proteins termed terpene synthases. The protein encoded by Lis is the first member of this family found to catalyze the formation of an acyclic monoterpene.
Enhanced levels of S-linalool by metabolic engineering of the terpenoid pathway in spike lavender leaves
DOI:10.1016/j.ymben.2014.03.003
PMID:24685653
[本文引用: 1]
Transgenic Lavandula latifolia plants overexpressing the linalool synthase (LIS) gene from Clarkia breweri, encoding the LIS enzyme that catalyzes the synthesis of linalool were generated. Most of these plants increased significantly their linalool content as compared to controls, especially in the youngest leaves, where a linalool increase up to a 1000% was observed. The phenotype of increased linalool content observed in young leaves was maintained in those T1 progenies that inherit the LIS transgene, although this phenotype was less evident in the flower essential oil. Cross-pollination of transgenic spike lavender plants allowed the generation of double transgenic plants containing the DXS (1-deoxy-d-xylulose-5-P synthase), coding for the first enzyme of the methyl-d-erythritol-4-phosphate pathway, and LIS genes. Both essential oil yield and linalool content in double DXS-LIS transgenic plants were lower than that of their parentals, which could be due to co-suppression effects linked to the structures of the constructs used. Copyright © 2014 International Metabolic Engineering Society. Published by Elsevier Inc. All rights reserved.
Enhanced levels of the aroma and flavor compound S-linalool by metabolic engineering of the terpenoid pathway in tomato fruits
The aromas of fruits, vegetables, and flowers are mixtures of volatile metabolites, often present in parts per billion levels or less. We show here that tomato (Lycopersicon esculentum Mill.) plants transgenic for a heterologous Clarkia breweri S-linalool synthase (LIS) gene, under the control of the tomato late-ripening-specific E8 promoter, synthesize and accumulate S-linalool and 8-hydroxylinalool in ripening fruits. Apart from the difference in volatiles, no other phenotypic alterations were noted, including the levels of other terpenoids such as gamma- and alpha-tocopherols, lycopene, beta-carotene, and lutein. Our studies indicate that it is possible to enhance the levels of monoterpenes in ripening fruits by metabolic engineering.
An unbiased approach elucidates variation in (S)-(+)-linalool,a context-specific mediator of a tri-trophic interaction in wild tobacco
Overexpression of LiTPS2 from a cultivar of lily (Lilium ‘Siberia’) enhances the monoterpenoids content in tobacco flowers
DOI:S0981-9428(20)30161-3
PMID:32278293
[本文引用: 1]
Lily, a famous cut flower with highly fragrance, has high ornamental and economic values. Monoterpenes are the main components contributing to its fragrance, and terpene synthase (TPS) genes play critical roles in the biosynthesis of monoterpenoids. To understand the function of TPS and to explore the molecular mechanism of floral scent in cultivar Lilium 'Siberia', transcriptomes of petal at different flowering stages and leaf were obtained by RNA sequencing and three unigenes related to TPS genes were selected for further validation. Quantitative real-time PCR showed that the expression level of LiTPS2 was greater than that of the other two TPS genes. Phylogenetic analysis indicated that LiTPS2 belonged to the TPSb subfamily, which was responsible for monoterpenes synthesis. Subcellular localization demonstrated that LiTPS2 was located in the chloroplasts. Furthermore, functional characterization showed that LiTPS2 utilized both geranyl pyrophosphate (GPP) and farnesyl pyrophosphate (FPP) to produce monoterpenoids such as linalool and sesquiterpenes like trans-nerolidol, respectively. Ectopic expression in transgenic tobacco plants suggested that the amount of linalool from the flowers of transgenic plants was 2-3 fold higher than that of wild-type plants. And the emissions of myrcene and (E)-β-ocimene were also accumulated from the flowers of LiTPS2 transgenic lines. Surprisingly, these three compounds were the main fragrance components of oriental lily hybrids. Our results indicated that LiTPS2 contributed to the production of monoterpenes and could effectively regulate the aroma of Lilium cultivars, laying the foundation for biotechnological modification of floral scent profiles.Copyright © 2020 Elsevier Masson SAS. All rights reserved.
Emission and accumulation of monoterpene and the key terpene synthase (TPS) associated with monoterpene biosynthesis in Osmanthus fragrans Lour
The terpene synthase gene family in Gossypium hirsutum harbors a linalool synthase GhTPS12 implicated in direct defence responses against herbivores
DOI:10.1111/pce.v41.1 URL [本文引用: 1]
Structure and evolution of linalool synthase
Plant terpene synthases constitute a group of evolutionarily related enzymes. Within this group, however, enzymes that employ two different catalytic mechanisms, and their associated unique domains, are known. We investigated the structure of the gene encoding linalool synthase (LIS), an enzyme that uses geranyl pyrophosphate as a substrate and catalyzes the formation of linalool, an acyclic monoterpene found in the floral scents of many plants. Although LIS employs one catalytic mechanism (exemplified by limonene synthase [LMS]), it has sequence motifs indicative of both LMS-type synthases and the terpene synthases employing a different mechanism (exemplified by copalyl diphosphate synthase [CPS]). Here, we report that LIS genes analyzed from several species encode proteins that have overall 40%-96% identity to each other and have 11 introns in identical positions. Only the region encoding roughly the last half of the LIS gene (exons 9-12) has a gene structure similar to that of the LMS-type genes. On the other hand, in the first part of the LIS gene (exons 1-8), LIS gene structure is essentially identical to that found in the first half of the gene encoding CPS. In addition, the level of similarity in the coding information of this region between the LIS and CPS genes is also significant, whereas the second half of the LIS protein is most similar to LMS-type synthases. Thus, LIS appears to be a composite gene which might have evolved from a recombination event between two different types of terpene synthases. The combined evolutionary mechanisms of duplication followed by divergence and/or "domain swapping" may explain the extraordinarily large diversity of proteins found in the plant terpene synthase family.
New perspectives in pollination biology:floral fragrances. A day in the life of a linalool molecule:chemical communication in a plant-pollinator system. Part 1:linalool biosynthesis in flowering plants
DOI:10.1046/j.1442-1984.1999.00014.x URL [本文引用: 1]
Diversity,regulation,and genetic manipulation of plant mono- and sesquiterpenoid biosynthesis
DOI:10.1007/s00018-009-0066-7 URL [本文引用: 1]
Characterization of S-(+)-linalool synthase from several provenances of Cinnamomum osmophloeum
DOI:10.1007/s11295-013-0665-1 URL [本文引用: 1]
Characterization of two monoterpene synthases involved in floral scent formation in Hedychium coronarium
DOI:10.1007/s00425-014-2127-x URL [本文引用: 1]
Identification and characterization of terpene synthase genes accounting for volatile terpene emissions in flowers of Freesia x hybrida
DOI:10.1093/jxb/ery224 URL [本文引用: 1]
Composition and biosynthesis of scent compounds from sterile flowers of an ornamental plant Clematis florida cv. ‘Kaiser’
DOI:10.3390/molecules25071711 URL [本文引用: 1]
The spatio-temporal expression of some genes involved in the biosynthetic pathways of terpenes/phenylpropanoids in yarrow (Achillea millefolium)
DOI:10.1016/j.plaphy.2019.06.036 URL
Essential oil production:relationship with abundance of glandular trichomes in aerial surface of plants
DOI:10.1007/s11738-008-0214-y URL [本文引用: 1]
Identification,functional characterization,and seasonal expression patterns of five sesquiterpene synthases in Liquidambar formosana
DOI:10.1021/acs.jnatprod.7b00773
PMID:29746128
[本文引用: 1]
Terpenoids are a large group of important secondary metabolites that are involved in a variety of physiological mechanisms, and many are used commercially in the cosmetics and pharmaceutical industries. During the past decade, the topic of seasonal variation in terpenoid biosynthesis has garnered increasing attention. Formosan sweet gum ( Liquidambar formosana Hance) is a deciduous tree species. The expression of terpene synthase and accumulation of terpenoids in leaves may vary in different seasons. Here, four sesquiterpene synthases (i.e., LfTPS01, LfTPS02, LfTPS03, and LfTPS04) and a bifunctional mono/sesquiterpene synthase ( LfTPS05) were identified from Formosan sweet gum. The gene expression of LfTPS01, LfTPS02, and LfTPS03 showed seasonal diversification, and, in addition, expression of LfTPS04 and LfTPS05 was induced by methyl jasmonate treatment. The major products LfTPS01, LfTPS02, LfTPS04, and LfTPS05 are hedycaryol, α-selinene, trans-β-caryophyllene, α-copaene/δ-cadinene, and nerolidol/linalool, respectively. The data indicated that the sesquiterpenoid content in the essential oil of Formosan sweet gum leaves shows seasonal differences that were correlated to the sesquiterpene synthase gene expression.
Linalool and linalool nerolidol synthases in roses,several genes for little scent
DOI:10.1016/j.plaphy.2018.03.009 URL [本文引用: 1]
Identification of a novel hedycaryol synthase gene isolated from Camellia brevistyla flowers and floral scent of Camellia cultivars
DOI:10.1007/s00425-015-2454-6 URL [本文引用: 1]
Codon usage and tRNA content in unicellular and multicellular organisms
Choices of synonymous codons in unicellular organisms are here reviewed, and differences in synonymous codon usages between Escherichia coli and the yeast Saccharomyces cerevisiae are attributed to differences in the actual populations of isoaccepting tRNAs. There exists a strong positive correlation between codon usage and tRNA content in both organisms, and the extent of this correlation relates to the protein production levels of individual genes. Codon-choice patterns are believed to have been well conserved during the course of evolution. Examination of silent substitutions and tRNA populations in Enterobacteriaceae revealed that the evolutionary constraint imposed by tRNA content on codon usage decelerated rather than accelerated the silent-substitution rate, at least insofar as pairs of taxonomically related organisms were examined. Codon-choice patterns of multicellular organisms are briefly reviewed, and diversity in G+C percentage at the third position of codons in vertebrate genes--as well as a possible causative factor in the production of this diversity--is discussed.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}