


水稻是三大主粮作物之一,我国水稻播种面积常年保持在3000万hm2,产量约2亿t[1]。淀粉是胚乳中含量最多的物质,其含量、组成成分及所占比例均会影响水稻的产量和品质。淀粉的合成是由多种酶共同参与的有序而复杂的生理生化过程,其中主要涉及的酶有5种,分别是ADP-葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase,AGPase)、可溶性淀粉合成酶(soluble starch synthase,SSS)、分支酶(branching enzyme,BE)、脱分支酶(debranching enzyme,DBE)以及颗粒结合淀粉酶(granule- bound starch synthase,GBSS)[2-3],以上酶功能的缺失均可能导致一系列异常水稻胚乳的产生。例如,AGPase由2个小亚基(small subunits,SSU)和4个大亚基(large subunits,LSU)构成,OsAGPL2负责编码AGPase的1个亚基,其发生突变会导致AGPase功能受损进而使胚乳产生皱缩表型[4];OsSSIIIa/Flo5编码淀粉合成酶IIIa(SSIIIa)蛋白,该基因突变致使SSIIIa功能异常,胚乳产生心白表型[5];淀粉分支酶(SBE)参与支链淀粉合成以及分支短链的形成,其功能缺陷时,导致支链淀粉链长变长、胚乳出现垩白和籽粒皱缩[6];异淀粉酶1(isoamylase 1,ISA 1)是一种DBE,当其失去原有的催化功能时,胚乳淀粉粒形成会受到阻碍,进而引起胚乳皱缩[7]。
间接影响淀粉合成相关酶的功能也会形成异常胚乳。例如,FLO6编码一个含有CBM48结构域的蛋白,与淀粉结合,并通过与SSIVb及GBSS相互作用调节淀粉合成,突变体flo6中的胚乳具有粉质不透明的表型,且胚乳内淀粉含量明显减少[8-9];FLO8编码UDP-葡萄糖焦磷酸化酶(UDP- glucose pyrophosphorylase 1,Ugp1),该基因突变使Ugp1活性降低,进而影响大部分淀粉合成酶的活性,导致胚乳粉质不透明、总淀粉和支链淀粉的含量降低、脂质含量升高、支链淀粉结构改变[10]。FLO2编码的蛋白含有3个可以介导蛋白质互作的四肽重复基序(TPR)结构域,在水稻胚乳发育过程中,FLO2蛋白可以调节一些与贮藏淀粉及贮藏蛋白合成有关基因的表达以及应对高温胁迫[11⇓-13],在水稻突变体flo2中,籽粒明显皱缩,胚乳粉质不透明,直链淀粉含量降低,支链淀粉结构改变[11]。
此外,与胚乳线粒体、造粉体和糊粉层发育、碳氮代谢、养分吸收转运相关的基因突变也会导致胚乳异常。基因FLO13负责编码线粒体复合体I附属亚基OsNDUFA9,其突变体的线粒体复合体I组装异常,导致线粒体结构改变、功能受损,使籽粒内干物质积累下降、复合淀粉粒排列疏松、胚乳粉质不透明[14];FLO10、Fse5、FLO18分别与线粒体NADH脱氢酶复合体I的3个亚基基因nad1、nad4和nad5的mRNA加工有关[15⇓-17]。FLO18负责nad5的5ʹ末端剪切,FLO10和Fse5分别负责nad1和nad4的第一内含子的反式剪切,它们的突变都会导致线粒体形态结构发生变化,胚乳表型异常。此外,FLO10对水稻糊粉层的发育也有影响。OsROS1与DNA去甲基化有关,其突变使DNA高甲基化以及可能与糊粉层发育有关的转录因子RISBZ1和RPBF表达受抑制,最终导致糊粉层的细胞层数增加,引起胚乳不透明、淀粉和直链淀粉含量减少、脂质和蛋白质含量提升[18]。FLO7影响造粉体发育及外围胚乳淀粉合成的功能,在突变体flo7中,外围胚乳呈现出粉质不透明的表型、胚乳内复合淀粉粒排列松散、淀粉组分及结构发生改变[19]。OsPPDKB编码丙酮酸磷酸双激酶(PPDKB),该基因与碳代谢有关,其突变体flo4籽粒不仅具有心白的表型,且胚乳中淀粉含量明显减少,脂肪酸含量升高[20]。OsBT1编码的蛋白与腺苷二磷酸葡萄糖(ADP-glucose,ADPglc)转运有关,其突变体由于OsBT1功能受损而使得在合成淀粉时ADPG供应不足,最终胚乳异常,总淀粉及直链淀粉的含量降低[21]。FLO19编码一个质体定位的丙酮酸脱氢酶复合物E1的亚基α1(ptPDC-E1-α1),该酶参与内半乳糖的生物合成,在突变体flo19中由于半乳糖合成减少而使造粉体发育异常、淀粉合成减少、复合淀粉颗粒形状异常、胚乳粉质不透明[22]。
本研究以一个来源于ZH11的粉质胚乳突变体cse-2为试验材料,通过对其农艺性状、籽粒表型和胚乳淀粉粒形态、胚乳组分含量、基因定位及测序验证等研究,发现突变体cse-2是flo2的1个新等位突变体。
1 材料与方法
1.1 试验材料
突变体cse-2源于粳稻品种ZH11(Oryza sativa L. ssp. japonica cv. Zhonghua 11)的组织培养突变体。突变体与籼稻品种IRAT129杂交获得F2定位群体,用于遗传分析及精细定位。以上材料于2021-2022年种植于安徽农业大学大杨镇基地,田间管理与大田生产一致。
1.2 测定指标与方法
1.2.1 农艺性状和遗传分析
成熟植株用于株高、分蘖数、节间长度、穗长、有效穗数、穗粒数、结实率等农艺性状的调查,利用t检验方法比较数据间差异性,用SPSS 20软件进行统计分析,用Excel软件进行数据处理并作图。
F1和F2代植株种子完全成熟后收获并脱壳,将糙米置于医用观片灯上观察胚乳透明情况,分析cse-2的遗传方式。
1.2.2 显微镜观察
用刀片横切突变体和野生型的籽粒,在体视镜下观察拍照。籽粒横切面经E1010型离子溅射仪(日立,日本)喷金镀膜处理后,利用S-4800型高分辨率扫描电子显微镜(日立,日本)观察并拍照[23]。
1.2.3 籽粒成分及含量
1.2.4 胚乳淀粉糊化温度
精米过100目筛后置于烘箱中35 ℃烘干备用。称取2.00 mg米粉样品,使用Pyris DSC 8000型差示扫描热量仪(DSC,Perten,美国)测定淀粉糊化温度以及糊化过程中的焓变[29]。所有试验均进行3次生物学重复。
1.3 突变基因定位及表达分析
采用图位克隆的方法对突变基因进行精细定位。提取F2叶片DNA[30],利用分布于水稻12条染色体上的79对具有多态性的分子标记对目的基因进行初步连锁分析。初步连锁后,利用
2 结果与分析
2.1 农艺性状及籽粒表型
图1

图1
野生型(ZH11)与突变体(cse-2)的表型比较
株高(a)、籽粒(b~e)。比例尺为10 cm(a)、1 cm(b~d)和1 mm(e)。
Fig.1
Comparison of phenotypes between wild type (ZH11) and mutant (cse-2)
Plant height (a) and grain (b-e). Bars=10 cm (a), 1 cm (b-d) and1 mm (e).
表1 野生型(ZH11)与突变体(cse-2)农艺性状比较
Table 1
| 农艺性状Agronomic trait | ZH11 | cse-2 |
|---|---|---|
| 株高Plant height (cm) | 104.05±0.56 | 94.55±1.18** |
| 主茎穗长Main panicle length (cm) | 23.80±0.46 | 21.22±0.51** |
| 第1节间长 The first internode length (cm) | 42.20±0.66 | 40.21±0.75 |
| 第2节间长 The second internode length (cm) | 19.16±0.64 | 18.57±0.40 |
| 第3节间长 The third internode length (cm) | 13.68±0.56 | 10.55±0.56** |
| 第4节间长 The fourth internode length (cm) | 7.35±0.14 | 5.24±0.17** |
| 有效分蘖数Effective tiller number | 20.75±2.47 | 24.85±2.56 |
| 一次枝梗数Primary branch number | 13.88±0.40 | 13.83±1.01 |
| 二次枝梗数Secondary branch number | 44.00±1.40 | 35.50±1.76** |
| 穗粒数Grain number per panicle | 221.85±6.84 | 191.90±6.11** |
| 结实率Seed-setting rate (%) | 89.84±0.13 | 80.58±0.19** |
| 粒长Grain length (mm) | 7.68±0.06 | 7.67±0.09 |
| 粒宽Grain width (mm) | 3.51±0.01 | 3.49±0.02 |
| 粒厚Grain thickness (mm) | 2.35±0.03 | 2.04±0.02** |
| 千粒重1000-kernel weight (g) | 25.63±0.40 | 19.16±0.17** |
“**”表示在P < 0.01水平上有极显著差异,下同。
“**”indicates extremely significant difference at P < 0.01 level, the same below.
图2

图2
野生型(ZH11)与突变体(cse-2)籽粒横截面扫描电镜图片
Fig.2
SEM images of grain cross section of wild type (ZH11) and mutant (cse-2)
2.2 突变体cse-2的遗传分析
突变体cse-2和籼稻IRAT129杂交后,F1胚乳均为正常表型,F2的胚乳表型发生了分离,其中籽粒胚乳表型正常的有191株,籽粒胚乳表型异常的有72株。经χ2检验符合孟德尔3:1分离比(χ2=0.67<χ20.05,1=3.84),表明突变体cse-2的表型是由1对隐性核基因控制。
2.3 籽粒理化性质分析
与野生型相比,突变体的总淀粉及直链淀粉含量分别极显著减少了22.4%和46.7%,而突变体蛋白质、脂肪酸及可溶性糖含量分别极显著增加了14.3%、78.4%、168.0%(图3)。
图3
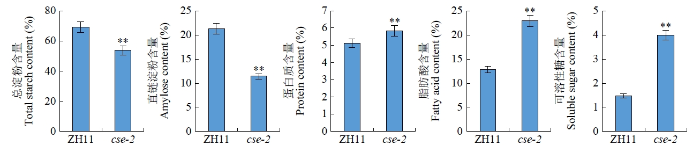
图3
野生型(ZH11)和突变体(cse-2)籽粒成分含量对比
“**”表示在P < 0.01水平上有极显著差异。
Fig.3
Comparison of grain component contents of wild type (ZH11) and mutant (cse-2)
“**”indicates extremely significant difference at P < 0.01 level.
差示扫描热量仪检测结果(表2)表明,突变体淀粉在糊化时的热特性与野生型间均存在极显著变化,其中突变体淀粉的糊化起始温度、峰值温度、终止温度分别下降4.7、4.1、2.5 ℃,糊化过程中吸收的热量显著减少10.8%。近红外快速分析仪测定的胶稠度以及碱消值均与野生型有极显著差异,其中突变体的碱消值极显著下降5.0%,而胶稠度极显著上升23.4%。以上结果表明,突变体淀粉的理化性质发生了明显变化。
表2 野生型(ZH11)与突变体(cse-2)大米理化性质比较
Table 2
| 材料 Material | 起始温度 To (℃) | 峰值温度 TP (℃) | 结束温度 TC (℃) | 焓变 ∆H (J/g) | 碱消值 Alkali consumption value | 胶稠度 Gel consistency (mm) |
|---|---|---|---|---|---|---|
| ZH11 | 66.90±0.55 | 72.20±0.37 | 76.10±0.27 | 6.50±0.22 | 8.10±0.37 | 56.40±0.18 |
| cse-2 | 62.20±0.31** | 68.10±0.48** | 73.60±0.17** | 5.80±0.11** | 7.70±0.48** | 69.60±0.15** |
2.4 CSE-2基因的精细定位及表达特性
图4
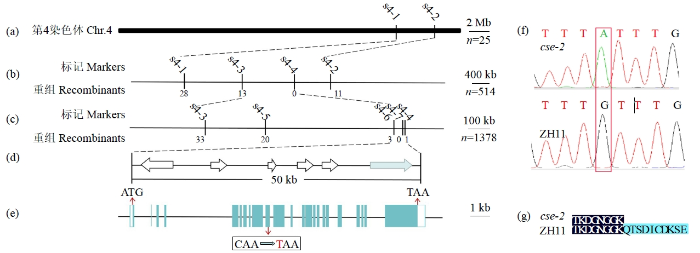
图4
CSE-2基因的精细定位
a~d:横线上方为定位所用的分子标记,横线下方为交换单株数,浅蓝色箭头表示候选基因;e:候选基因CSE-2结构图,浅蓝色框代表外显子,其中间空白部分代表内含子,红色箭头分别表示起始密码子、终止密码子、突变位点,黑线框内容表示突变位点;f:目的基因反向测序峰图,红线框内为突变位点;g:部分蛋白质序列比对,无义突变引起的氨基酸缺失。
Fig.4
Fine mapping of CSE-2 gene
a-d: the numbers above and beneath the bold lines represent the molecular markers and recombinants, respectively, and the light blue arrow represent the candidate gene; e: the structure diagram of candidate gene CSE-2, the light blue box represent the exon, the blank part represent the intron, the red arrows represent the start codon, stop codon and mutation site, respectively, and the content in the black box represent the mutation; f: target gene reverse sequencing peak map, with mutated sites in the red line box; g: partial protein sequence alignment near the none-sense mutation site.
图5

图5
野生型(ZH11)与突变体(cse-2)中FLO2蛋白质序列比对
红色箭头表示cse-2突变体内FLO2突变形成终止密码子的位点。
Fig.5
Sequence alignment of FLO2 protein between wild type (ZH11) and mutant (cse-2)
Red arrow indicates the site that the stop codon occurred of FLO2 in the cse-2 mutant.
3 讨论
基因LOC-Os04g55230曾被报道并命名为FLO2[11],目前已有多个flo2的等位突变体被报道,She等[11]通过诱变得到8个突变位点位于第11(2个突变位点)、14(3个突变位点一样)、21外显子以及第14和21内含子的剪切位点上flo2的等位突变体;Qiao等[31]借助诱变手段获得了3个flo2等位突变体,其突变位点分别位于第1、14(2个)外显子上;Wu等[32]通过诱变获得了3个flo2的等位突变体,其突变位点分别位于第9、11内含子的3′ 剪切位点以及第19外显子;邱家静[33]通过60Co辐射诱变得到9个flo2的等位突变体,其突变位点分别位于第13、14(2个)、16和17内含子剪切位点以及第6、10(编码区1431位点突变)、11、21外显子处。本研究鉴定了一个具有粉质皱缩胚乳的水稻突变体cse-2,是由基因LOC-Os04g55230第6064位碱基C突变成T引起无义突变导致的,该突变位点与以往发现的FLO2突变位点不同,故cse-2是FLO2的一个新等位基因,突变体是flo2的一个新等位突变体。FLO2蛋白含有介导蛋白互作的TPR结构域及一个功能不明确的CLU结构域,由于cse-2中该基因的无义突变,使翻译的蛋白只含有N端的CLU结构域。一些参与多肽运输、控制分蘖及花粉育性有关的蛋白质同样含有TPR结构域[34-35],这表明FLO2除影响贮藏淀粉及贮藏蛋白的合成外,可能还具有其他生理功能,对于cse-2的研究将为TPR结构域的功能研究提供基因资源。
cse-2的胚乳外围表型正常,而中心部分粉质不透明,这与She等[11]、Qiao等[31]、邱家静[33]报道的表型类似,与Wu等[32]报道的3个暗胚突变体不同。此外,而本研究中的cse-2的粒长比野生型明显降低,这与邱家静[33]报道的同为第10外显子的等位突变体M227的糙米粒长表型不同。突变体cse-2的株高、主茎穗长、穗粒数和结实率等均有显著变化,而突变体flo2在这些农艺性状方面与野生型并无差异,这可能是由于2种突变体内FLO2蛋白序列不同引起的。cse-2胚乳中总淀粉和直链淀粉含量明显降低,蛋白质、脂肪酸以及可溶性糖含量却极显著升高,这表明由于淀粉合成途径受阻,而对多种初级和次级代谢产物的产生影响,导致脂肪酸等中间产物合成积累增加。此外,有报道[36]指出,CSE-2在拟南芥中的同源基因AtFLO2的突变引起脂质合成有关基因的表达水平降低,进而导致胚乳内脂肪酸含量下降,这一结果与cse-2中的情况相反,表明CSE-2及其同源基因在不同物种中对合成脂肪酸的影响机制可能有所不同。cse-2胚乳与报道过的胚乳突变体w24[4]、flo5[6]、fse5[18]、flo6[10]和osagpl2-3[37]等均为胚乳中心粉质不透明。但这些突变体籽粒内的淀粉粒形态相似,osagpl2-3的淀粉粒虽然大小不一但形状也一致[37],而cse-2内部的淀粉粒大小不一且形态多样。
4 结论
cse-2胚乳具有粉质不透明的表型、胚乳内的淀粉粒排列松散、淀粉粒外形异常,并且在株高、穗粒数、结实率等农艺性状方面与野生型存在显著差异。此外,突变体的淀粉含量显著低于野生型,蛋白质、脂肪酸以及可溶性糖的含量高于野生型。在理化性质上,突变体淀粉的热特性指数显著低于野生型,表明突变体淀粉更易糊化。遗传分析表明该性状受1对核隐性基因控制,图位克隆以及测序结果表明,基因LOC_Os04g55230的一个碱基替换导致该突变性状,该基因曾被报道为FLO2,故突变体cse-2是flo2的一个新的等位突变体。
参考文献
中国水稻生产、市场与进出口贸易的回顾与展望
DOI:10.3969/j.issn.1006-8082.2021.04.005
[本文引用: 1]

新中国成立以来,我国水稻科技不断取得重大突破,单产连创新高,年人均稻谷占有量突破150 kg,稻米供求实现了由长期短缺到总量平衡有余的历史性跨越,优质稻占比持续提高;市场从统购统销到合同定购,从保护价收购到托市收购,市场化程度不断加深,国家宏观调控能力不断增强,稻米市场价格稳步上涨;国家持续加强对大米贸易的宏观调控,改革开放以来的大多数年份里我国都是大米净出口国,是国际市场重要的大米供应商,但2011年以来进口大米数量快速增加,大米逐步呈现净进口。长期看,我国水稻生产、市场、贸易等方面仍然存在一些突出问题,需要加快培育推广优质高产专用水稻品种,推广绿色高质高效生产技术,促进水稻产业绿色高质量发展。
The complexities of starch biosynthesis in cereal endosperms
DOI:10.1016/j.copbio.2008.02.013
PMID:18400487
[本文引用: 1]
Starch serves not only as an energy source for plants, animals, and humans but also as an environmentally friendly alternative for fossil fuels. Here, we describe recent findings concerning the synthesis of this important molecule in the cereal endosperm. Results from six separate transgenic reports point to the importance of adenosine diphosphate glucose pyrophosphorylase in controlling the amount of starch synthesized. The unexpected cause underlying the contrast in sequence divergence of its two subunits is also described. A major unresolved question concerning the synthesis of starch is the origin of nonrandom or clustered alpha-1,6 branch-points within the major component of starch, amylopectin. Developing evidence that several of the starch biosynthetic enzymes involved in amylopectin synthesis occur in complexes is reviewed. These complexes may provide the specificity for the formation of nonrandom branch-points.
Formation of starch in plant cells
ADP-glucose pyrophosphorylase large subunit 2 is essential for storage substance accumulation and subunit interactions in rice endosperm
Knockout of a starch synthase gene OsSSIIIa/Flo5 causes white-core floury endosperm in rice (Oryza sativa L.)
Starch-branching enzyme I-deficient mutation specifically affects the structure and properties of starch in rice endosperm
DOI:10.1104/pp.103.021527
PMID:14526120
[本文引用: 2]
We have isolated a starch mutant that was deficient in starch-branching enzyme I (BEI) from the endosperm mutant stocks of rice (Oryza sativa) induced by the treatment of fertilized egg cells with N-methyl-N-nitrosourea. The deficiency of BEI in this mutant was controlled by a single recessive gene, tentatively designated as starch-branching enzyme mutant 1 (sbe1). The mutant endosperm exhibited the normal phenotype and contained the same amount of starch as the wild type. However, the mutation apparently altered the fine structure of amylopectin. The mutant amylopectin was characterized by significant decrease in both long chains with degree of polymerization (DP) > or = 37 and short chains with DP 12 to 21, marked increase in short chains with DP < or = 10 (A chains), and slight increase in intermediate chains with DP 24 to 34, suggesting that BEI specifically synthesizes B1 and B2-3 chains. The endosperm starch from the sbe1 mutant had a lower onset concentration for urea gelatinization and a lower onset temperature for thermo-gelatinization compared with the wild type, indicating that the genetic modification of amylopectin fine structure is responsible for changes in physicochemical properties of sbe1 starch.
Endosperm sugar accumulation caused by mutation of PHS 8/ISA 1 leads to pre-harvest sprouting in rice
FLOURY ENDOSPERM6 encodes a CBM48 domain-containing protein involved in compound granule formation and starch synthesis in rice endosperm
The CBM48 domain-containing protein FLO 6 regulates starch synthesis by interacting with SSIVb and GBSS in rice
FLOURY ENDOSPERM8, encoding the UDP-glucose pyrophosphorylase 1, affects the synthesis and structure of starch in rice endosperm
A novel factor FLOURY ENDOSPERM2 is involved in regulation of rice grain size and starch quality
Overexpression of rice OsLEA5 relieves the deterioration in seed quality caused by high-temperature stress
Reduced rice grain production under high-temperature stress closely correlates with ATP shortage during seed development
OsNDUFA9 encoding a mitochondrial complex I subunit is essential for embryo development and starch synthesis in rice
Rice FLOURY ENDOSPERM10 encodes a pentatricopeptide repeat protein that is essential for the trans-splicing of mitochondrial nad1 intron 1 and endosperm development
Rice FLOURY SHRUNKEN ENDOSPERM 5 encodes a putative plant organelle RNA recognition protein that is required for cis-splicing of mitochondrial nad4 intron 1
Rice FLOURY ENDOSPERM 18 encodes a pentatricopeptide repeat protein required for 5′processing of mitochondrial nad 5 messenger RNA and endosperm development
Mutations in the DNA demethylase OsROS1 result in a thickened aleurone and improved nutritional value in rice grains
FLOURY ENDOSPERM7 encodes a regulator of starch synthesis and amyloplast development essential for peripheral endosperm development in rice
DOI:10.1093/jxb/erv469
PMID:26608643
[本文引用: 1]
In cereal crops, starch synthesis and storage depend mainly on a specialized class of plastids, termed amyloplasts. Despite the importance of starch, the molecular machinery regulating starch synthesis and amyloplast development remains largely unknown. Here, we report the characterization of the rice (Oryza sativa) floury endosperm7 (flo7) mutant, which develops a floury-white endosperm only in the periphery and not in the inner portion. Consistent with the phenotypic alternation in flo7 endosperm, the flo7 mutant had reduced amylose content and seriously disrupted amylopectin structure only in the peripheral endosperm. Notably, flo7 peripheral endosperm cells showed obvious defects in compound starch grain development. Map-based cloning of FLO7 revealed that it encodes a protein of unknown function. FLO7 harbors an N-terminal transit peptide capable of targeting functional FLO7 fused to green fluorescent protein to amyloplast stroma in developing endosperm cells, and a domain of unknown function 1338 (DUF1338) that is highly conserved in green plants. Furthermore, our combined β-glucuronidase activity and RNA in situ hybridization assays showed that the FLO7 gene was expressed ubiquitously but exhibited a specific expression in the endosperm periphery. Moreover, a set of in vivo experiments demonstrated that the missing 32 aa in the flo7 mutant protein are essential for the stable accumulation of FLO7 in the endosperm. Together, our findings identify FLO7 as a unique plant regulator required for starch synthesis and amyloplast development within the peripheral endosperm and provide new insights into the spatial regulation of endosperm development in rice. © The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology.
White-core endosperm floury endosperm-4 in rice is generated by knockout mutations in the C4-type pyruvate orthophosphate dikinase gene (OsPPDKB)
Analysis of the rice ADP-glucose transporter (OsBT1) indicates the presence of regulatory processes in the amyloplast stroma that control ADP-glucose flux into starch
DOI:10.1104/pp.15.01911
PMID:26754668
[本文引用: 1]
Previous studies showed that efforts to further elevate starch synthesis in rice (Oryza sativa) seeds overproducing ADP-glucose (ADPglc) were prevented by processes downstream of ADPglc synthesis. Here, we identified the major ADPglc transporter by studying the shrunken3 locus of the EM1093 rice line, which harbors a mutation in the BRITTLE1 (BT1) adenylate transporter (OsBt1) gene. Despite containing elevated ADPglc levels (approximately 10-fold) compared with the wild-type, EM1093 grains are small and shriveled due to the reduction in the amounts and size of starch granules. Increases in ADPglc levels in EM1093 were due to their poor uptake of ADP-[(14)C]glc by amyloplasts. To assess the potential role of BT1 as a rate-determining step in starch biosynthesis, the maize ZmBt1 gene was overexpressed in the wild-type and the GlgC (CS8) transgenic line expressing a bacterial glgC-TM gene. ADPglc transport assays indicated that transgenic lines expressing ZmBT1 alone or combined with GlgC exhibited higher rates of transport (approximately 2-fold), with the GlgC (CS8) and GlgC/ZmBT1 (CS8/AT5) lines showing elevated ADPglc levels in amyloplasts. These increases, however, did not lead to further enhancement in seed weights even when these plant lines were grown under elevated CO2. Overall, our results indicate that rice lines with enhanced ADPglc synthesis and import into amyloplasts reveal additional barriers within the stroma that restrict maximum carbon flow into starch. © 2016 American Society of Plant Biologists. All Rights Reserved.
Plastidic pyruvate dehydrogenase complex E 1 component subunit Alpha1 is involved in galactolipid biosynthesis required for amyloplast development in rice
GPA5 encodes a Rab5a effector required for post-Golgi trafficking of rice storage proteins
Development of NIRS models to predict protein and amylose content of brown rice and proximate compositions of rice bran
Predicting single kernel and bulk milled rice alkali spreading value and gelatinization temperature class using NIR spectroscopy
Effects of variations in the chemical composition of individual rice grains on the eating quality of hybrid indica rice based on near-infrared spectroscopy
Rapid isolation of high molecular weight plant DNA
DOI:10.1093/nar/8.19.4321
PMID:7433111
[本文引用: 1]
A method is presented for the rapid isolation of high molecular weight plant DNA (50,000 base pairs or more in length) which is free of contaminants which interfere with complete digestion by restriction endonucleases. The procedure yields total cellular DNA (i.e. nuclear, chloroplast, and mitochondrial DNA). The technique is ideal for the rapid isolation of small amounts of DNA from many different species and is also useful for large scale isolations.
Fine mapping and candidate gene analysis of the floury endosperm gene, FLO(a), in rice
Three novel alleles of FLOURY ENDOSPERM2 (FLO2) confer dull grains with low amylose content in rice
TPR domain coding gene ST2 may be involved in regulating tillering and fertility in rice
Evolutionarily evolved discriminators in the 3-TPR domain of the Toc64 family involved in protein translocation at the outer membrane of chloroplasts and mitochondria
Arabidopsis thaliana FLO2 is involved in efficiency of photoassimilate translocation,which is associated with leaf growth and aging,yield of seeds and seed quality
DOI:10.1093/pcp/pcw217
PMID:28158741
[本文引用: 1]
FLO2, FLOURY ENDOSPERM 2, is highly conserved in higher plants, and rice FLO2 has been predicted to be involved in regulation of accumulation of storage compounds. We analyzed the function of Arabidopsis thaliana FLO2 (AtFLO2) because A. thaliana set structurally different seeds from those of rice. Although the flo2 mutant of A. thaliana showed normal germination, inflorescence and morphogenesis of flowers, peculiar phenotypes on leaves and siliques were observed, suggesting that this gene played important roles during both the vegetative and reproductive stages. The mutant leaves showed a decrease in chloroplast numbers, and increased total biomass with faster growth. When grown in high light intensity conditions, it was observed that aging events were induced. The flo2 mutant showed depressed transportation of photoassimilates into the sink organs. In the reproductive stage, the flo2 mutant had significantly smaller size siliques, causing a reduced yield of seeds. These seeds were structurally weak, and the quality of seeds was significantly lowered, with reduction of accumulation of storage compounds by seeds. A positron-emitting tracer imaging system (PETIS) analysis detected a decreased amount of photoassimilate transport in the flo2 mutant. Therefore, it was presumed that the phenotypes of the flo2 mutant were caused by reduced performance of translocation or transportation of the photoassimilates. Our observation suggests that AtFLO2 is strongly involved in regulation of translocation and transport of assimilates, and contributes greatly to quality control of the various processes involving substance supply or transfer, such as photoassimilation, leaf enlargement, yield of seeds in a silique and accumulation of seed storage compounds.© The Author 2017. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Phenotypic and candidate gene analysis of a new floury endosperm mutant (osagpl2-3) in rice
Comparative study on the structure-properties relationships of native and debranched rice starch
Quality of different rice cultivars and factors influencing consumer willingness-to-purchase rice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}