


专性寄生菌咖啡驼孢锈菌(Hemileia vastatrix)引起的真菌性病害是威胁全球咖啡安全生产的毁灭性病害[1]。云南省是我国最重要的咖啡生产区,截至2021年,全省咖啡种植面积9.29万hm2,全国占比超过98%,几乎全省咖啡种植区均不同程度受叶锈病的危害[2]。该病可引起咖啡植株衰弱,叶片过早脱落,形成枝枯[3],严重时落叶率高达50%以上,导致咖啡减产30%~50%,甚至出现绝产[4]。近年来,为提高精品咖啡占比,铁毕卡(Typica)、波邦(Buorbon)、瑰夏(Geisha)等易感锈病品种开始广泛种植[5]。由于卡蒂姆(Catimor)系列品种抗锈性衰退[6]及叶锈病病原菌生理小种变异等[7],叶锈病对云南省咖啡安全生产已构成了严重威胁。光合作用作为植物干物质积累的唯一途径[8],在病害侵染后会破坏植物叶片光系统,导致叶绿素含量降低,进而光合量子产量降低、电子传递链受阻、光合速率下降,最终导致产量降低、品质变劣[9-10]。但已有研究[11]表明,病害胁迫下,植物的净光合速率(Pn)、胞间CO2摩尔分数(Ci)及气孔导度(Gs)呈先降低后升高的规律,叶绿素a/b的含量出现短暂上升。由此表明,在病害胁迫下植物间光合特征的变化特点不一致。小粒咖啡(Coffea arabica)属茜草科(Rubiaceae)咖啡属(Coffea)植物,国内关于小粒咖啡光合特征的研究主要涉及低温胁迫[12]、不同遮阴环境[13]、间套作[14]、不同品种[15]及海拔梯度[16]对光合特征的影响,而关于咖啡光合特征对叶锈病侵染胁迫的响应机制尚不明确。叶锈病是制约咖啡生长和产量稳定的关键病害,开展叶锈病对咖啡光合特征影响的研究对明确咖啡的抗病机理和科学防控具有指导意义。而植物受真菌、细菌等病原菌侵染时,体内防御酶活性也会发生一系列变化[17]。其中,过氧化物酶(POD)、超氧化物歧化酶(SOD)、几丁质酶(CHT)及β-1, 3-葡聚糖酶(β-1, 3-GA)是植物中研究最多、特征最明确的防御相关性蛋白[17-18]。POD、SOD、CHT及β-1, 3-GA作为植物体内重要的抗氧化酶和细胞保护酶系统,与植物抗病性密切相关[16,18]。其中,CHT及β-1, 3-GA很早便被证实参与植物对真菌病原体的抗性,两者协同可降解真菌细胞壁,具有水解作用,能够遏制多种病原真菌的生长[16,19]。目前,关于4种抗逆性酶在不结球白菜霜霉病[11]、大麦叶斑病[16]及花生黑腐病[18]等多种病原菌侵染后的活性变化做了较为系统的研究。叶锈病是一种世界性真菌病害,但关于其病原菌咖啡驼孢锈菌侵染后4种抗逆性酶活性的变化规律尚不明确。鉴于此,本研究以0~Ⅶ共8个叶锈病危害等级(或咖啡叶锈病感染面积)的咖啡叶片为研究对象,对不同危害等级下咖啡的落叶率(LF)、叶绿素相对含量(SPAD)、光合特征及4种抗逆性酶活性进行测定,以期明确叶锈病不同危害等级下咖啡叶片落叶、SPAD、光合特征及抗逆性酶活性的变化规律,为进一步研究咖啡叶锈病的精准防控及其真菌与寄主植物互作提供数据支撑。
1 材料与方法
1.1 试验地概况
试验于2023年在云南省保山市佐园咖啡有限公司瑰夏种植基地(98°50′26″ E,25°29′59″ N,海拔1247±5 m)进行。该咖啡基地始建于2017年,面积约26.67 hm2,坡度不低于25°,阴坡种植,黑色沙壤土。种植咖啡品种为Geisha,采用等高线开台种植,株距1.0~1.5 m,行距超过3 m。咖啡平均株高1.6 m,平均冠幅2.6 m,平均地径64.14 mm,采用咖啡单一种植模式。咖啡园于2022年采用黑色防草布进行全园覆盖除草,并建设有标准的水肥一体化设施。该区域为典型干热河谷型气候类型,年均温17.25 ℃,最低月均温10 ℃,最高月均温22 ℃,全年无霜,月均降水量在74 mm左右,全年旱雨季分明,降水主要集中于7月和8月。
1.2 测定指标与方法
1.2.1 落叶率
图1

图1
叶锈病危害等级划分标准
Fig.1
Classification standard for the harm level of leaf rust disease
1.2.2 叶绿素含量
2023年10月初,随机选取8个危害等级下的咖啡叶片,每个危害等级选取15枚,共计135枚。通过SPAD-502叶绿素仪(日本Konica公司)测定选取叶片的SPAD。叶片选取标准为一级分枝上从顶芽向下的第3或第4对当年成熟的咖啡叶片。
1.2.3 光合参数
同期,在晴天无云的上午9:00- 11:00使用Li-6400便携式光合仪(美国LI-COR公司)对Pn、Gs、Ci、蒸腾速率(Tr)及光合有效辐射(PAR)进行测定。测定过程中8个危害等级下各随机选择15枚叶片,叶片选取标准与SPAD测定的一致。并计算各危害等级下咖啡叶片的水分利用率(EWU)和光能利用率(LUE)[15]。EWU=Pn/Tr;LUE=Pn/PAR。
1.2.4 抗逆性酶
同期,随机选择受叶锈病侵染的15株咖啡植株,每株分别随机选取8个危害等级下咖啡叶片5枚,叶片选取标准为与SPAD测定的一致。5株为1个重复,共3个重复,每个重复下将5株咖啡植株8个危害等级下采集的叶片样品分别混合后,使用锡箔纸包裹,并用棉线缠绕记录后,置于充满液氮的15 L液氮罐内临时保存。带回实验室后转移至超低温冰箱保存。测定时使用液氮对样品进行充分研磨。按照试剂盒(苏州格瑞生物科技有限公司,江苏)的说明书对POD、SOD、CHT及β-1, 3-GA 4种抗逆性酶进行提取和活性测定。
1.3 数据处理
使用Excel 2020软件进行数据整理,使用SPSS 19.0、Origin 2020和Photoshop 2021软件进行数据分析和绘图。
2 结果与分析
2.1 不同危害等级下落叶率比较分析
由图2可以看出,不同危害等级下咖啡落叶率有差异,Ⅲ~Ⅶ共5个危害等级落叶率显著高于危害等级0~Ⅱ,并且5个危害等级彼此间均具有显著性差异,危害等级Ⅶ的落叶率最高,为33.95%,排序为Ⅲ<Ⅳ<Ⅴ<Ⅵ<Ⅶ;叶锈病感染面积0.00%~2.00%间落叶率较低,0~Ⅱ共3个危害等级落叶率差异不显著。整体来看,落叶率随着叶锈病感染面积的增加而增加。
图2
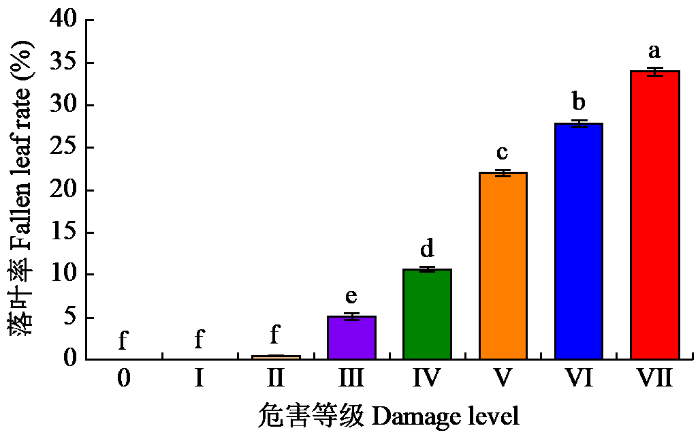
图2
不同危害等级下咖啡落叶率
不同小写字母表示差异显著(P < 0.05)。下同。
Fig.2
Coffee fallen leaf rate under different damage levels
Different lowercase letters indicate significant difference at P < 0.05. The same below.
2.2 不同危害等级下SPAD比较分析
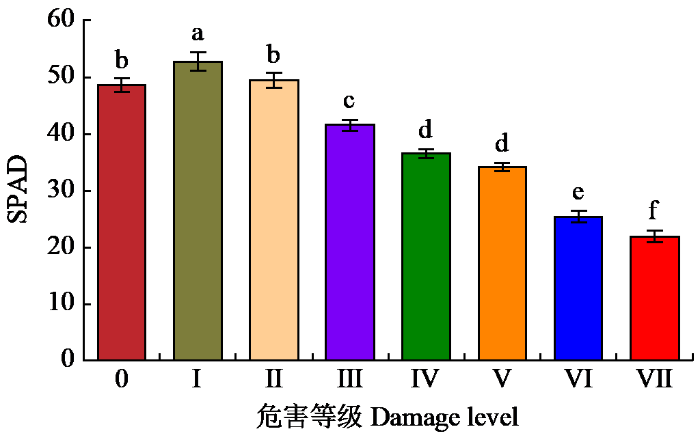
由图3可以看出,危害等级Ⅰ的SPAD最高;危害等级0与Ⅱ的SPAD次之,彼此差异不显著,但均显著高于危害等级Ⅲ~Ⅶ,显著低于危害等级Ⅰ;危害等级Ⅲ~Ⅶ的SPAD逐级递减,除危害等级Ⅳ和Ⅴ彼此间差异不显著,其余危害等级下彼此间均具有显著性差异,危害等级Ⅶ的最低。
图3
2.3 不同危害等级下光合参数比较分析
由表1可知,咖啡叶片危害等级Ⅲ的Pn显著高于除危害等级Ⅰ外的其他6个危害等级,危害等级Ⅶ显著低于其他7个危害等级,其余6个危害等级的Pn排序为Ⅰ>Ⅱ>Ⅳ>Ⅴ>0>Ⅵ。危害等级Ⅲ的Gs显著高于其他7个危害等级的,危害等级0的显著低于其他7个危害等级的,危害等级Ⅰ~Ⅱ与危害等级Ⅳ~Ⅶ共6个危害等级间差异不显著。危害等级Ⅶ的Ci显著高于其余7个危害等级的,危害等级Ⅶ与Ⅳ、Ⅲ彼此间差异不显著,但显著高于0~Ⅱ和Ⅴ共4个危害等级的;0~Ⅴ共6个危害等级间差异不显著。咖啡叶片危害等级Ⅲ的Tr显著高于其余7个危害等级的,危害等级0的显著低于其余7个危害等级的,而Ⅰ~Ⅱ和Ⅳ~Ⅶ共6个危害等级的Tr彼此间差异不显著。咖啡叶片危害等级0~Ⅰ间的EWU差异不显著,但危害等级0显著高于其他6个危害等级;危害等级Ⅰ与Ⅱ和Ⅲ间差异不显著,但显著高于Ⅳ和Ⅴ,而危害等级Ⅱ~Ⅴ间差异不显著;危害等级Ⅶ与Ⅵ差异不显著,但显著低于其他6个危害等级;咖啡叶片危害等级Ⅳ的LUE最高,与危害等级Ⅱ和Ⅲ的差异不显著;危害等级0~Ⅲ和Ⅴ~Ⅵ共6个危害等级间差异不显著;危害等级Ⅶ的最低,但与0~Ⅱ和Ⅴ~Ⅵ共5个危害等级间差异不显著。
表1 不同危害等级下咖啡叶片的光合参数
Table 1
| 危害等级 Damage level | Pn [μmol/(m2·s)] | Gs [mol/(m2·s)] | Ci (μmol/mol) | Tr [mmol/(m2·s)] | EWU (mmol/mol) | LUE (mol/mmol) |
|---|---|---|---|---|---|---|
| 0 | 9.50±1.88cd | 0.08±0.01c | 230.90±27.27c | 1.43±0.23c | 6.82±1.28a | 1.14±0.26bc |
| Ⅰ | 15.58±2.14ab | 0.14±0.03b | 223.78±21.09c | 3.17±0.53b | 5.31±0.55ab | 1.51±0.17bc |
| Ⅱ | 14.75±2.14bc | 0.16±0.02b | 243.00±27.88c | 3.73±0.49b | 4.34±0.72bc | 1.56±0.33abc |
| Ⅲ | 20.65±3.48a | 0.23±0.03a | 267.44±27.04bc | 5.56±0.48a | 4.02±0.78bc | 1.88±0.24ab |
| Ⅳ | 11.74±2.13bc | 0.14±0.01b | 273.60±19.55bc | 3.96±0.40b | 2.89±0.44cd | 3.33±1.62a |
| Ⅴ | 11.49±1.51bcd | 0.13±0.02b | 252.70±21.44c | 3.77±0.46b | 3.20±0.45cd | 1.14±0.21bc |
| Ⅵ | 6.08±1.16d | 0.13±0.02b | 326.44±13.05b | 4.23±0.46b | 1.52±0.25de | 0.48±0.10bc |
| Ⅶ | 0.20±0.10e | 0.13±0.01b | 438.00±12.22a | 4.15±0.37b | 0.05±0.03e | 0.02±0.01c |
同列数据后不同小写字母表示处理间在P < 0.05水平差异显著。
Different lowercase letters after the data in the same column indicate significant differences between treatments at P < 0.05 level.
2.4 不同危害等级下抗逆性酶比较分析
2.4.1 SOD活性变化
由图4可知,危害等级0~Ⅵ的SOD活性变化不明显,介于45.50~313.87 U/(g·h) FW,危害等级Ⅶ最高,为2686.41 U/(g·h) FW。整体来看,SOD活性随着咖啡叶锈病感染面积的增加呈平缓变化后急剧升高,叶锈病感染前期对SOD活性的影响较小。
图4

2.4.2 POD活性变化
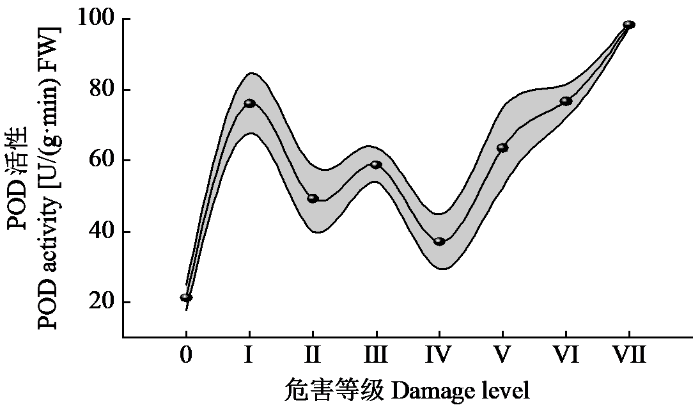
由图5可知,危害等级0的POD活性最低,为21.33 U/(g·min) FW;危害等级Ⅰ的POD活性急剧升高,为76.17 U/(g·min) FW;危害等级Ⅱ~Ⅵ的POD活性波动较大,但均高于危害等级0;危害等级Ⅵ以后POD活性呈上升趋势,危害等级Ⅶ达到最大值,为98.50 U/(g·min) FW。整体来看,受咖啡叶锈病感染后,均能一定程度提高POD活性。
图5
2.4.3 CHT活性变化
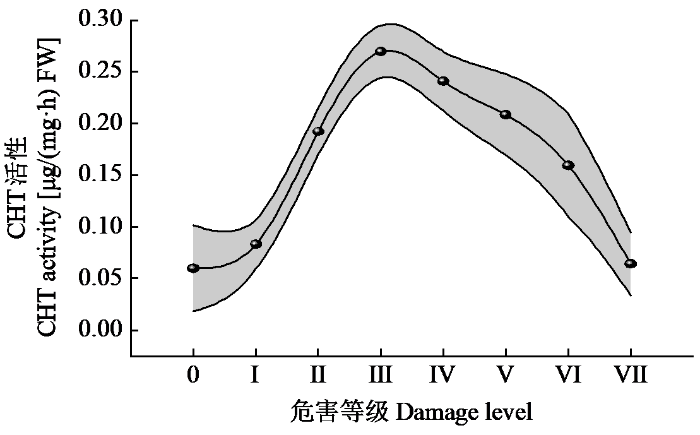
由图6可知,随着危害等级的增加CHT活性先快速升高后又逐渐降低,危害等级Ⅲ的CHT活性达最大,为0.27 μg/(mg·h) FW。
图6
2.4.4 β-1, 3-GA活性变化
由图7可知,危害等级0~Ⅶ的β-1, 3-GA活性变化与CHT活性变化基本一致,呈先快速升高后又逐渐降低,在危害等级Ⅲ下活性达到最大值,为125.50 mg/(min·g) FW。
图7

2.5 咖啡叶片光合作用与抗逆性酶各指标的相关性分析
由图8可知,不同危害等级下咖啡叶片光合作用与抗逆性酶指标间具有显著相关性,其中显著正相关和显著负相关各8对。CHT活性与β-1, 3-GA活性、SOD活性与Ci、β-1, 3-GA活性与Pn和Gs、Pn与SPAD、Gs与Tr、Ci与LF、EWU与SPAD呈显著正相关,相关系数分别为0.81、0.90、0.75、0.83、0.73、0.86、0.85和0.92;SOD活性与Pn、POD活性与EWU、Pn与Ci和LF、Ci与EWU和SPAD、EWU与LF、SPAD与LF呈显著负相关,相关系数分别为-0.74、-0.72、-0.78、-0.77、-0.88、-0.86、-0.91和-0.97。
图8
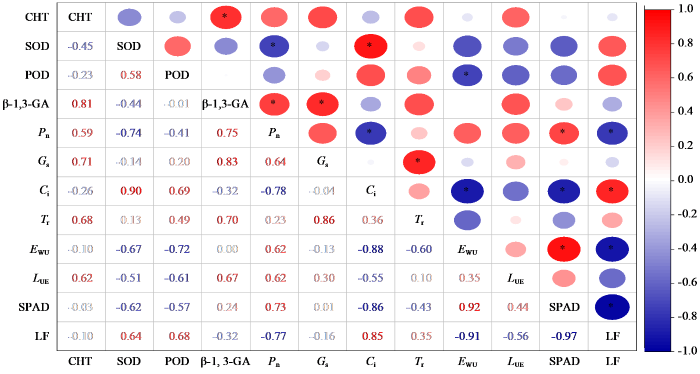
图8
咖啡叶片光合作用与抗逆性酶各指标的相关性分析
“*”表示达显著相关水平(P < 0.05)。
Fig.8
Correlation analysis between photosynthesis and various indicators of stress resistant enzymes in coffee leaves
“*”indicates significant correlation at P < 0.05 level.
2.6 不同危害等级下咖啡叶片综合评价分析
由表2可知,不同危害等级下咖啡叶片综合得分排序为Ⅲ>Ⅱ>Ⅳ>Ⅰ>Ⅴ>0>Ⅵ>Ⅶ,危害等级Ⅲ的综合得分大于1,为1.94。结果表明危害等级Ⅲ下咖啡叶片光合作用更强、抗逆性更高、落叶相对较少。
表2 不同危害等级下咖啡叶片的综合得分
Table 2
| 危害等级 Damage level | 主成分得分 Score of principal components | 综合得分 Comprehensive score | 排序 Rank | |
|---|---|---|---|---|
| PC1 | PC2 | |||
| 0 | 1.17 | -3.99 | -0.65 | 6 |
| Ⅰ | 1.56 | -0.85 | 0.53 | 4 |
| Ⅱ | 1.73 | 0.16 | 0.96 | 2 |
| Ⅲ | 1.93 | 2.94 | 1.94 | 1 |
| Ⅳ | 1.19 | 1.03 | 0.95 | 3 |
| Ⅴ | -0.21 | 0.20 | -0.04 | 5 |
| Ⅵ | -2.11 | 0.47 | -0.95 | 7 |
| Ⅶ | -5.23 | 0.03 | -2.74 | 8 |
3 讨论
植物落叶受气候条件、病虫害、品种特性、管理水平等多种因素影响[21]。叶锈病作为小粒咖啡的关键性病害,发生严重时可导致超过50%的叶片脱落[4]。本次调查发现,受叶锈病侵染后咖啡叶片会脱落,该结果与前人[4]研究一致。但本次调查还发现落叶率与叶锈病侵染面积的大小关系密切,即叶锈病侵染面积低于4.01%时,仅会引起少落叶,甚至不落叶,但随着叶锈病侵染面积增加,落叶率也显著增加,超过90%以上的落叶是在侵染面积超过4.01%时发生的,原因可能是随着叶锈病侵染面积增加,尤其是危害中后期,锈斑上产生大量的锈孢子,导致叶绿素含量降低,叶绿体被破坏,从而引起叶片大量脱落。而在SPAD测定中也发现,随着叶锈病侵染面积增加SPAD出现短暂上升后急剧下降的情况。
叶绿素作为植物光合色素之一,在光合作用过程中发挥吸收、传递及转化光能的功能[22]。通常植物受病害侵染后叶绿素含量呈下降趋势[22]。整体来看,在受叶锈病侵染后SPAD整体呈下降趋势,但侵染面积0.10%~0.99%时SPAD被显著提高。已有研究[23]证实,病害侵染后导致叶绿素含量降低的原因主要是受病害侵染后叶片发黄或溃烂所致。而叶锈病侵染初期并不会导致叶片黄化等症状,但随着危害程度增加病斑扩大,病斑上形成大量的淡黄色锈孢子,尤其是中后期病斑出现干枯,呈褐色,导致SPAD降低。叶锈病侵染初期,也有可能是叶片通过增加叶绿素含量来增强光能转化为化学能适应或应答叶锈病胁迫,在相关性分析中也证实了SPAD与Pn呈显著正相关关系,表明叶片确实通过增加叶绿素含量来提高光能转化率应对叶锈病胁迫。
光合作用作为植物生长发育过程中最重要的生理过程之一,可以将光能转化为植物体内能够贮藏利用的化学能[10]。在病害胁迫下植物的光合作用会受影响,如锈病、霜霉病、白粉病等真菌性病害,其病原菌通过形成霉层,导致光合作用的部位和面积减少,进而影响光能吸收、传递和转化、电子传递及光合磷酸化,最终严重制约光合生产[24]。本次调查结果表明,叶锈病侵染可显著影响咖啡的光合作用,主要表现为Pn、Gs、Tr和LUE均随着侵染面积的增加先增加后降低,而Ci则持续增加,EWU则持续降低。随着病害侵染面积增加,Gs显著增加,从而导致大量的CO2进入,而到病害侵染后期因危害导致叶片内部细胞空隙加大,增加了CO2的进入空间,导致Ci随病害发病程度呈增加趋势。其次,气孔活动受限制时,蒸腾作用也会受限制[23]。本研究表明,在受叶锈病侵染后Gs并没有受限制,侵染后均大于正常叶片,这也是Tr被提高的关键原因,而从相关分析中也证实了Gs与蒸腾作用具有显著正相关关系。研究[23]表明,较高的蒸腾作用有利于根部无机营养的快速转运,从而间接刺激了光合作用。而本次调查表明在危害等级Ⅲ下蒸腾作用最强,也表明该时期叶片的无机营养转运能力最强。病害后期因叶片组织被严重损害,已失去自我防御能力,叶片内部细胞空隙增加,增加了CO2进入空间,导致Ci随病害发生严重程度呈缓慢增加后急剧增加,出现危害等级Ⅶ下Ci最大的情况。Ci作为评判气孔限制和非气孔限制的重要依据,Pn、Gs及Ci同时下降时,光合作用为气孔限制,反之Pn随着Ci增加而降低,说明光合作用限制因素为非气孔限制[23]。本研究结果表明,危害等级0~Ⅲ时,Pn、Gs及Ci同时增加,表明该时期主要为气孔限制;而危害等级Ⅲ以后的Pn随着Ci的增加而降低,表明该时期为非气孔限制。
叶锈病侵染后,并没有立即导致光合速率降低,而是在危害等级Ⅳ后才开始下降,表明该时期后咖啡叶片光能转化为化学能的机能开始降低,随着侵染面积的增加,最直观的就是从危害等级Ⅳ后,90%以上的落叶发生在该时期之后,在整个侵染期咖啡叶片SOD、POD、CH和β-1, 3-GA的活性也发生了变化。SOD和POD作为抗氧化酶,在提高植物抗逆性中发挥重要作用[25]。本研究结果表明,在咖啡叶锈病侵染后SOD和POD 2种抗氧化酶活性均在危害等级Ⅶ下(咖啡叶锈病感染面积20.01%~50.00%)达到最高水平,这可能与该时期产生大量活性氧相关,有待进一步进行测定验证。此外,2种酶前期变化规律并不一致,这主要与POD不仅与活性氧清除和逆境适应性相关,还参与酚类物质氧化,抑制褐变,并在调节植物生长发育、木质素合成中发挥重要作用。植物产生的CHT和β-1, 3-GA对真菌性病害具有防卫反应,两者协同降解真菌细胞壁起水解酶作用,可强烈抑制真菌的生长,植物受病害胁迫后2种酶的活性均升高[26],该结果已在葡萄霜霉病[27]、柑橘褐斑病[28]、大麦叶斑病[16]中得到证实。本研究结果表明,咖啡叶片在受叶锈病侵染后CHT和β-1, 3-GA的活性均呈上升趋势,该结果与上述病害的研究结果一致。表明受专性寄生菌咖啡驼孢锈菌侵染后能够诱导咖啡叶片内CHT和β-1, 3-GA的活性明显增高,以抑制病原菌的生长。值得关注的是,上述2种酶均在危害等级Ⅲ下(咖啡叶锈病感染面积2.01~4.00%)活性达到峰值,也表明了该时期具有更高的抗病性,随着咖啡叶锈病侵染面积的增加抗锈逐渐衰减。同时,相关分析表明CHT与β-1, 3-GA的活性呈显著正相关关系,从而进一步证实了2种酶降解病原真菌细胞壁具有协同作用。
通过主成分分析方法进行综合评价分析,结果显示,危害等级Ⅲ(咖啡叶锈病感染面积2.01%~ 4.00%)下综合得分最高,为1.94%,表明该时期咖啡叶片光合作用更强,抗逆性更高,落叶相对较少,对咖啡生长影响较低。过去咖啡叶锈病防控主要以化学防控为主[29],多在病害发生早期进行防控。本研究结果表明,在叶锈病感染面积低于4.01%时,叶片的光合作用并不受影响,且植物自身具有较强的抗性,并不需要开展化学防控,建议通过灌溉、施肥等农业管理措施增强植物自身抗性,减少化学农药的使用。
4 结论
叶锈病侵染后会导致叶片脱落,落叶率随着叶锈病侵染面积的增加而逐渐增加,叶锈病侵染面积低于4.01%时仅会引起少量落叶,甚至不落叶;90%以上的落叶是在侵染面积超过4.01%时发生的。SPAD随着叶锈病侵染面积的增加先短暂上升后急剧降低,初期,叶锈病侵染面积低于2.01%时,可以一定程度提高SPAD。在叶锈病侵染面积介于2.01%~4.00%时,叶片的Pn、Gs、Tr及Ci最大,EWU随着叶锈病侵染面积的增加而降低,EUE则先升高后降低,叶锈病侵染后显著降低了EWU;在侵染面积低于8.01%时可一定程度提高LWU,表明叶锈病侵染后对咖啡叶片光合产生了显著性影响。同时,CHT和β-1, 3-GA的活性随着叶锈病侵染面积增加呈先增加后降低的规律;SOD活性则随着咖啡呈先平缓变化后急剧升高;POD活性则波动较大,但在侵染后活性均提高,表明4种酶均在咖啡叶锈病侵染后作出了响应。综合分析表明,叶锈病危害等级Ⅲ(咖啡叶锈病感染面积2.01%~4.00%)下叶片光合作用更强,抗逆性更高,落叶也相对较少。因此,在叶锈病侵染面积低于4.01%时,不建议开展化学防控,建议通过灌溉、施肥等农业管理措施增强植物自身抗性,减少化学农药的使用。
参考文献
2021年度云南省咖啡产业发展报告
(2022-07-09) [2024-02-27]. https://nync.yn.gov.cn/uploadfile/s38/2022/0811/20220811110742328.pdf.
Becoming ʻWildʼ at the intersection of knowledges: Coffee rust crisis in Costa Rica
Elucidating the interactions between the rust Hemileia vastatrix and a Calonectria mycoparasite and the coffee plant
Photosynthetic decline in flag leaves of two field-grown spring wheat cultivars with different senescence properties
Effects of pyraclostrobin on leaf diseases, leaf physiology, yield and quality of durum wheat under Mediterranean conditions
白粉病对小麦光合特性的影响及病害严重度的定量模拟
DOI:10.3864/j.issn.0578-1752.2023.06.004
[本文引用: 2]

【目的】明确白粉病胁迫对小麦光合特性的影响规律,构建白粉病胁迫下小麦光合生产的模拟模型。【方法】以小麦为试验材料,分别于拔节期和孕穗期进行不同接种程度的小麦白粉病试验,明确白粉病对小麦光合特性的影响规律;在此基础上构建小麦白粉病严重度预测模型,量化白粉病对小麦的生理影响;基于单叶净光合速率(P<sub>n</sub>)和叶面积指数(LAI),实现小麦白粉病严重度预测模型与作物生长模型(WheatGrow)的耦合。【结果】白粉病胁迫下,小麦单叶P<sub>n</sub>和LAI均呈现下降趋势,与对照(CK)相比分别平均下降18.81%和23.41%,且与初始接种程度相比,发病时期对小麦P<sub>n</sub>和LAI的影响更为严重;小麦白粉病田间病情发展具有明显的平缓期、指数爆发期和稳定期,总的来说各处理下小麦白粉病流行的时间动态变化特征符合Logistic函数,基于白粉病胁迫对小麦影响的拟合结果,构建小麦白粉病病害胁迫因子,用以反映白粉病对小麦生理指标影响的胁迫效应;基于WheatGrow模型的光合生产子模型,结合小麦白粉病病害胁迫因子,提出模拟白粉病对小麦叶片P<sub>n</sub>和LAI效应的算法,并利用独立年份的数据资料对改进后的WheatGrow模型进行检验。【结论】耦合白粉病胁迫因子的WheatGrow模型对白粉病胁迫下小麦叶片P<sub>n</sub>、LAI、地上部生物量和产量的预测精度均好于原模型,模拟精度较原模型分别提高了53.29%、43.61%、60.09%和67.57%,改进后的模型可为小麦白粉病严重度的预测与小麦产量损失的定量评估等提供数字化工具和技术支撑。
β-1,3-glucanase and chitinase activities in soybean root nodules
Induction of β-1, 3- glucanase in seedlings of pearl millet in response to infection by sclerospora graminicola
Standard area diagram with color photographs to estimate the severity of coffee leaf rust in Coffea canephora
Antifungal hydrolases in pea tissue: II: inhibition of fungal growth by combinations of chitinase and β-1, 3-glucanase
DOI:10.1104/pp.88.3.936
PMID:16666407
[本文引用: 1]
Chitinase and beta-1,3-glucanase purified from pea pods acted synergistically in the degradation of fungal cell walls. The antifungal potential of the two enzymes was studied directly by adding protein preparations to paper discs placed on agar plates containing germinated fungal spores. Protein extracts from pea pods infected with Fusarium solani f.sp. phaseoli, which contained high activities of chitinase and beta-1,3-glucanase, inhibited growth of 15 out of 18 fungi tested. Protein extracts from uninfected pea pods, which contained low activities of chitinase and beta-1,3-glucanase, did not inhibit fungal growth. Purified chitinase and beta-1,3-glucanase, tested individually, did not inhibit growth of most of the test fungi. Only Trichoderma viride was inhibited by chitinase alone, and only Fusarium solani f.sp. pisi was inhibited by beta-1,3-glucanase alone. However, combinations of purified chitinase and beta-1,3-glucanase inhibited all fungi tested as effectively as crude protein extracts containing the same enzyme activities. The pea pathogen, Fusarium solani f.sp. pisi, and the nonpathogen of peas, Fusarium solani f.sp. phaseoli, were similarly strongly inhibited by chitinase and beta-1,3-glucanase, indicating that the differential pathogenicity of the two fungi is not due to differential sensitivity to the pea enzymes. Inhibition of fungal growth was caused by the lysis of the hyphal tips.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}