
作物杂志,2022, 第6期: 42–53 doi: 10.16035/j.issn.1001-7283.2022.06.006
OsWD40过表达水稻根系响应盐胁迫的转录组分析
闻丹妮( ), 鲍聆然, 刘蒙蒙, 沈波()
), 鲍聆然, 刘蒙蒙, 沈波()
- 杭州师范大学生命与环境科学学院,311121,浙江杭州
Transcriptome Analysis of OsWD40 Overexpression Rice Roots in Response to Salt Stress
Wen Danni(), Bao Lingran, Liu Mengmeng, Shen Bo()
- College of Life and Environmental Sciences, Hangzhou Normal University, Hangzhou 311121, Zhejiang, China
摘要:
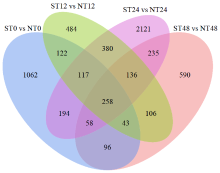
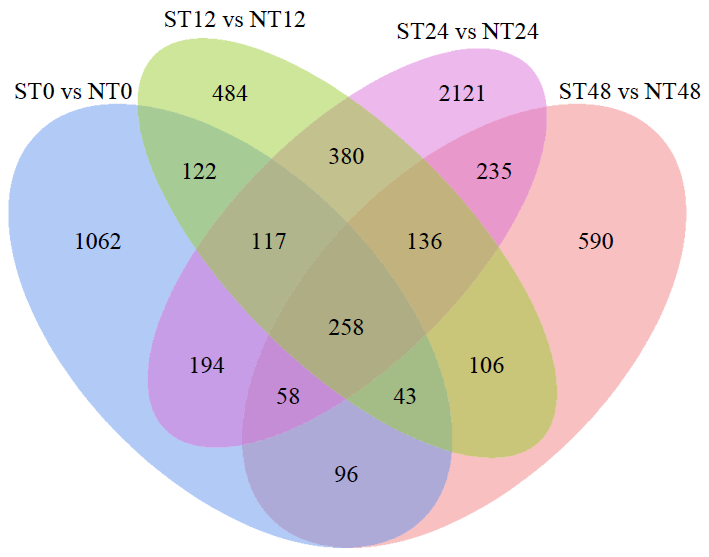
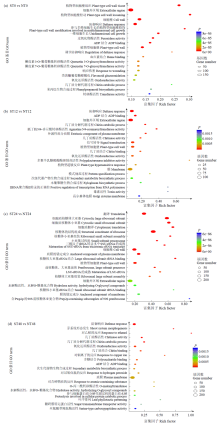
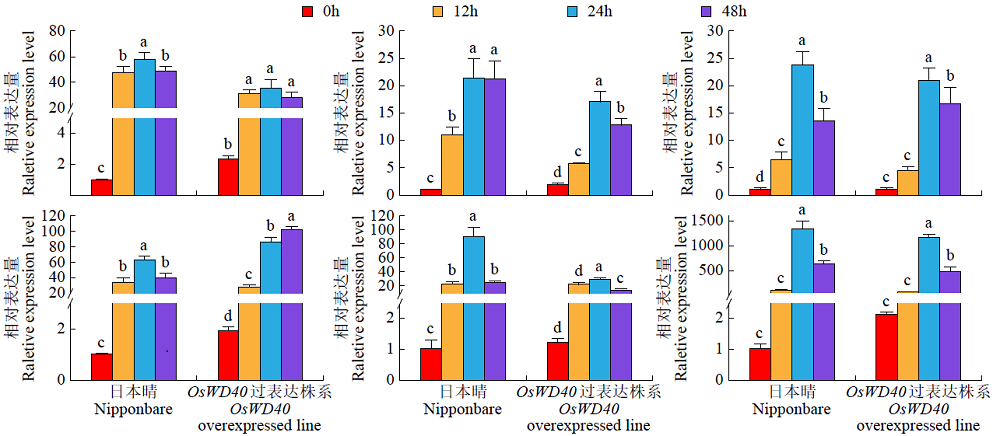
盐胁迫是影响水稻产量的主要因素之一,开展水稻耐盐机制的研究十分必要。为了揭示OsWD40基因参与耐盐的分子机制,以日本晴和OsWD40过表达水稻株系为材料,用浓度为200mmol/L的NaCl分别处理0、12、24和48h,对其根系进行转录组测序分析。结果显示,比较日本晴和OsWD40过表达株系在盐胁迫相同时间(ST0与NT0、ST12与NT12、ST24与NT24、ST48与NT48)的基因表达量,分别检测到1950、1646、3499和1522个差异表达基因。其中,盐胁迫处理24h的差异表达基因多于0、12和48h处理。对4个比较组的差异表达基因分别进行GO功能富集分析和KEGG代谢通路分析,发现差异表达基因主要富集在盐胁迫响应、脱落酸响应和转录调控等GO条目中,富集的重要代谢通路主要是植物激素信号转导,植物MAPK信号传导途径和苯丙烷生物合成、类黄酮生物合成相关的次生代谢途径等。同时,转录因子家族基因,如WRKY、MYB和bHLH等,在各比较组中呈现差异表达。由此推测,苯丙烷生物合成和类黄酮生物合成等植物次生代谢途径在OsWD40过表达水稻根系响应盐胁迫中发挥着重要作用,而且OsWD40可能介导响应脱落酸的基因转录调控,激活下游盐胁迫相关基因的表达。





| [1] |
Almeida D M, Oliveira M M, Saibo N J M. Regulation of Na+ and K+ homeostasis in plants:towards improved salt stress tolerance in crop plants. Genetics and Molecular Biology, 2017, 40(1):326-345.
doi: 10.1590/1678-4685-gmb-2016-0106 |
| [2] |
Senadheera P, Singh R, Maathuis F J. Differentially expressed membrane transporters in rice roots may contribute to cultivar dependent salt tolerance. Journal of Experimental Botany, 2009, 60(9):2553-2563.
doi: 10.1093/jxb/erp099 pmid: 19395386 |
| [3] |
Negrao S, Schmockel S M, Tester M, et al. Evaluating physiological responses of plants to salinity stress. Annals of Botany, 2017, 119:1-11.
doi: 10.1093/aob/mcw191 pmid: 27707746 |
| [4] |
梁永书, 周军杰, 南文斌, 等. 水稻根系研究进展. 植物学报, 2016, 51(1):98-106.
doi: 10.11983/CBB15011 |
| [5] |
Stirnimann C U, Petsalaki E, Russell R B, et al. WD40 proteins propel cellular networks. Trends in Biochemical Sciences, 2010, 35(10):565-574.
doi: 10.1016/j.tibs.2010.04.003 pmid: 20451393 |
| [6] |
Smith T F. Diversity of WD-repeat proteins. Sub-Cellular Biochemistry, 2008, 48:20-30.
doi: 10.1007/978-0-387-09595-0_3 pmid: 18925368 |
| [7] |
Xu C, Min J. Structure and function of WD40 domain proteins. Protein Cell, 2011, 2(3):202-214.
doi: 10.1007/s13238-011-1018-1 pmid: 21468892 |
| [8] |
Smith T F, Gaitatzes C, Saxena K, et al. The WD repeat:a common architecture for diverse functions. Trends in Biochemical Sciences, 1999, 24(5):181-185.
pmid: 10322433 |
| [9] | Nocker S V, Ludwig P. The WD-repeat protein superfamily in Arabidopsis:conservation and divergence in structure and function. BMC Genomics, 2003, 4(1):297-300. |
| [10] |
Xu X Z, Wan W, Jiang G B, et al. Nucleocytoplasmic trafficking of the Arabidopsis WD 40 repeat protein XIW1 regulates ABI5 stability and abscisic acid responses. Molecular Plant, 2019, 12(12):1598-1611.
doi: 10.1016/j.molp.2019.07.001 |
| [11] |
Kong D J, Li M J, Dong Z H, et al. Identification of TaWD40D,a wheat WD40 repeat-containing protein that is associated with plant tolerance to abiotic stresses. Plant Cell Reports, 2015, 34:395-410.
doi: 10.1007/s00299-014-1717-1 |
| [12] |
Huang J, Wang M M, Bao Y M, et al. SRWD:A novel WD 40 protein subfamily regulated by salt stress in rice (Oryza sativa L.). Gene, 2008, 424(1):71-79.
doi: 10.1016/j.gene.2008.07.027 |
| [13] |
Wang J, Zhang Y D, Fan F J, et al. Comparative transcriptome analysis reveals molecular response to salinity stress of salt-tolerant and sensitive genotypes of indica rice at seedling stage. Scientific Reports, 2018, 8(1):1755-1767.
doi: 10.1038/s41598-018-20191-w |
| [14] |
Chen E Y, Shen B. OsABT,a rice WD40 domain-containing protein,is involved in abiotic stress tolerance. Rice Science, 2022, 29(3):247-256.
doi: 10.1016/j.rsci.2021.07.012 |
| [15] |
Wang A J, Shu X Y, Jing X, et al. Identification of rice (Oryza sativa L.) genes involved in sheath blight resistance via a genome-wide association study. Plant Biotechnology Journal, 2021, 19(8):1553-1566.
doi: 10.1111/pbi.13569 |
| [16] |
Horie T, Sugawara M, Okada T, et al. Rice sodium-insensitive potassium transporter,OsHAK5,confers increased salt tolerance in tobacco BY2 cells. Journal of Bioscience and Bioengineering, 2011, 111(3):346-356.
doi: 10.1016/j.jbiosc.2010.10.014 |
| [17] |
Yang T, Feng H, Zhang S, et al. The potassium transporter OsHAK 5 alters rice architecture via ATP-dependent transmembrane auxin fluxes. Plant Communications, 2020, 1(5):100052.
doi: 10.1016/j.xplc.2020.100052 |
| [18] |
Sah S K, Reddy K R, Li J X. Abscisic acid and abiotic stress tolerance in crop plants. Frontiers in Plant Science, 2016, 7:571.
doi: 10.3389/fpls.2016.00571 pmid: 27200044 |
| [19] |
Nambara E, Marion P A. Abscisic acid biosynthesis and catabolism. Annual Review of Plant Biology, 2005, 56:165-185.
pmid: 15862093 |
| [20] |
Kim H, Lee K, Hwang H, et al. Overexpression of PYL5 in rice enhances drought tolerance,inhibits growth,and modulates gene expression. Journal of Experimental Botany, 2014, 65(2):453-464.
doi: 10.1093/jxb/ert397 |
| [21] |
Zhou M J, Guan Y H, Ren H B, et al. A bZIP transcription factor,OsABI5,is involved in rice fertility and stress tolerance. Plant Molecular Biology, 2008, 66(6):675-683.
doi: 10.1007/s11103-008-9298-4 |
| [22] | 孙立影, 于志晶, 李海云. 植物次生代谢物研究进展. 吉林农业科学, 2009, 34(4):4-10. |
| [23] |
Fraser C M, Chaoole C. The Phenylpropanoid Pathway in Arabidopsis. The Arabidopsis Book, 2011, 9:e0152.
doi: 10.1199/tab.0152 |
| [24] |
Gui J S, Shen J H, Li L G. Functional characterization of evolutionarily divergent 4-coumarate:coenzyme a ligases in rice. Plant Physiology, 2011, 157(2):574-586.
doi: 10.1104/pp.111.178301 pmid: 21807887 |
| [25] | 崔慧萍, 周薇, 郭长虹. 植物过氧化物酶体在活性氧信号网络中的作用. 中国生物化学与分子生物学报, 2017, 33(3):220-226. |
| [26] |
Huang J, Sun S, Xu D, et al. A TFIIIA-type zinc finger protein confers multiple abiotic stress tolerances in transgenic rice (Oryza sativa L.). Plant Molecular Biology, 2012, 80(3):337-350.
doi: 10.1007/s11103-012-9955-5 pmid: 22930448 |
| [27] |
Devaiah B N, Karthikeyan A S, Raghothama K G. WRKY75 transcription factor is a modulator of phosphate acquisition and root development in Arabidopsis. Plant Physiology, 2007, 143(4):1789-1801.
doi: 10.1104/pp.106.093971 |
| [28] | Pillai S E, Kumar C, Patel H K, et al. Overexpression of a cell wall damage induced transcription factor,OsWRKY42,leads to enhanced callose deposition and tolerance to salt stress but does not enhance tolerance to bacterial infection. BioMed Central, 2018, 18(1):177. |
| [29] | Zhao Y, Xing L, Wang X A, et al. The ABA receptor PYL 8 promotes lateral root growth by enhancing MYB77-dependent transcription of auxin-responsive genes. Science Signaling, 2014, 7(328):ra53. |
| [30] |
Dai X Y, Xu Y Y, Ma Q B, et al. Overexpression of an R1R2R3 MYB gene,OsMYB3R-2,increases tolerance to freezing,drought,and salt stress in transgenic Arabidopsis. Plant Physiology, 2007, 143(4):1739-1751.
doi: 10.1104/pp.106.094532 |
| [31] |
Liu G Z, Li X L, Jin S X, et al. Overexpression of rice NAC gene SNAC1 improves drought and salt tolerance by enhancing root development and reducing transpiration rate in transgenic cotton. PLoS ONE, 2017, 9(1):e86895.
doi: 10.1371/journal.pone.0086895 |
| [32] |
Schiefelbein J, Huang L, Zheng X H. Regulation of epidermal cell fate in Arabidopsis roots:the importance of multiple feedback loops. Frontiers in Plant Science, 2014, 5:47.
doi: 10.3389/fpls.2014.00047 pmid: 24596575 |
| [33] |
Li J Z, Han Y C, Lei L, et al. qRT9,a quantitative trait locus controlling root thickness and root length in upland rice. Journal of Experimental Botany, 2015, 66(9):2723-2732.
doi: 10.1093/jxb/erv076 |
| [34] |
Cao Y F, Song F M, Goodman R M, et al. Molecular characterization of four rice genes encoding ethylene-responsive transcriptional factors and their expressions in response to biotic and abiotic stress. Journal of Plant Physiology, 2006, 163(11):1167-1178.
pmid: 16436304 |
| [1] | 周浩, 邱先进, 徐建龙. 磁化水灌溉对农作物生长发育影响的研究进展[J]. 作物杂志, 2022, (6): 1–6 |
| [2] | 秦猛, 崔士泽, 何孝东, 翟玲侠, 陶博, 王召君, 赵海成, 李红宇, 郑桂萍, 刘丽华. 秸秆膨化还田对水稻产量、品质及土壤养分的影响[J]. 作物杂志, 2022, (6): 159–166 |
| [3] | 姜树坤, 王立志, 杨贤莉, 张喜娟, 刘凯, 迟力勇, 李锐, 来永才. 1961-2019年松嫩平原盐碱地区域水稻生长季气候资源的时空变化特征分析[J]. 作物杂志, 2022, (6): 214–219 |
| [4] | 种浩天, 尚程, 张运波, 黄礼英. 增密减氮对不同类型水稻品种颖花形成的影响[J]. 作物杂志, 2022, (6): 226–233 |
| [5] | 王金香, 王艳芝, 幸丽璇, 刘建霞, 王润梅. 赤霉素对盐胁迫下绿宝糯黍子幼苗根生长及渗透调节的影响[J]. 作物杂志, 2022, (6): 98–104 |
| [6] | 王瀚祥, 李广存, 徐建飞, 王万兴, 金黎平. 植物耐盐机理研究进展[J]. 作物杂志, 2022, (5): 1–12 |
| [7] | 王燕, 李廷友, 王豆, 李佳薇, 彭雯璐, 芮海云. 异甜菊醇对盐胁迫下小麦幼苗生长的影响[J]. 作物杂志, 2022, (5): 141–145 |
| [8] | 李睿, 董立强, 商文奇, 于广星, 代贵金, 王铮, 李跃东. 水稻苗期不同喷淋间隔处理对其生长发育及产量的影响[J]. 作物杂志, 2022, (5): 249–254 |
| [9] | 董林林, 沈明星, 施林林, 沈园, 王海候, 陆长婴. 生物质炭配施蚯蚓粪对水稻产量及养分吸收的影响[J]. 作物杂志, 2022, (5): 69–77 |
| [10] | 周宇娇, 张伟杨, 杨建昌. 高温胁迫导致水稻光温敏核不育系开颖与雌蕊受精障碍的研究进展[J]. 作物杂志, 2022, (4): 1–8 |
| [11] | 陈士勇, 王锐, 陈志青, 张海鹏, 王娟娟, 单玉华, 杨艳菊. 纳米锌和离子锌对水稻产量形成及籽粒锌含量的影响[J]. 作物杂志, 2022, (4): 107–114 |
| [12] | 郑思怡, 杨晔, 宋远辉, 花芹, 林泉祥, 张海涛, 程治军. 水稻甜质胚乳突变体m5788的鉴定及基因定位[J]. 作物杂志, 2022, (4): 14–21 |
| [13] | 王元元, 谷子寒, 陈平平, 易镇邪. 镉污染稻田玉米对水稻的季节性替代种植可行性研究[J]. 作物杂志, 2022, (4): 187–192 |
| [14] | 张海鹏, 陈志青, 王锐, 卢豪, 崔培媛, 杨艳菊, 张洪程. 氮肥配施纳米镁对水稻产量、品质和氮肥利用率的影响[J]. 作物杂志, 2022, (4): 255–261 |
| [15] | 施娴, 李洪有, 卢丙越, 周云, 赵继菊, 赵孟丽, 梁京, 孟衡玲. 3个苦荞品种对盐胁迫的生理响应及耐受性评价[J]. 作物杂志, 2022, (3): 149–154 |
|
||