


农杆菌(Agrobacterium tumefaciens)凭借其天然的T-DNA递送系统,已成为植物遗传转化的核心工具[1
近年来,针对宿主限制的机制研究揭示了植物―农杆菌互作的关键调控节点。例如,植物内源水杨酸(SA)可通过抑制VirA激酶活性负向调控Vir基因表达[10],而外源添加乙酰香草酸可拮抗SA的抑制作用[11]。在此基础上,载体系统设计取得显著进展:超级双元载体通过组成型启动子驱动VirE1/VirE2持续表达以增强T复合物稳定性[12],而三元载体系统通过辅助质粒补充VirG等关键调控元件,使难转化玉米品种的转化效率提升3~5倍[13-14]。然而,现有改良策略多聚焦于质粒元件优化,作为遗传操作核心的农杆菌基因组工程研究却长期停滞。实验室常用菌株(如EHA105)仍沿用20世纪80年代的遗传背景,其基因组缺乏适配现代基因编辑技术的工程化改造,难以满足精准调控与稳定性需求。
1 材料与方法
1.1 试验材料
供试农杆菌菌株为EHA105感受态细胞(ZC142-2,北京庄盟国际生物基因科技有限公司),大肠杆菌菌株为Trans1-T1(CD501-03,北京全式金生物技术有限公司),抗生素(卡那霉素储存液100 mg/mL、利福平储存液50 mg/mL和壮观霉素储存液50 mg/mL),载体(pCAMBIA3301基础载体),同源重组酶NEBuilder HiFi DNA Assembly Master Mix(E2621L,北京百灵克生物科技有限责任公司),Phanta SE Super-Fidelity DNA Polymerase(P521-d2,南京诺唯赞生物科技股份有限公司),DNA损伤试剂甲基磺酸甲酯(CM7242-5g,北京酷来搏科技有限公司)。
1.2 试验方法
1.2.1 碱基编辑载体构建
ABE碱基编辑载体以本实验室保存的pCAMBIA3301为骨架载体进行构建,构建内容主要包括:(1)由细菌组成型启动子pLac启动的dCas9(D10A,H840A)分别融合TadA8e、TadA8.20和TadA8r脱氨酶的蛋白表达框,3种脱氨酶分别连接在dCas9(D10A,H840A)的N端,以T3T terminator为终止子。(2)esgRNA由细菌强组成型启动子pJ23119启动,以rrnB T2 terminator终止。(3)sgRNA靶标序列来自A. tumefaciens EHA105的recA基因(5’-TCCCAGA TCGAACGGTCGTT-3’)。在远离PAM端的5和7 bp处各有1个A碱基作为ABE碱基编辑器的测试靶点。
CBE碱基编辑载体以本实验室保存的pCAMBIA3301基因编辑基础载体为骨架载体进行构建,构建内容主要包括:(1)由细菌组成型启动子pLac或农杆菌诱导型启动子pvirB启动的dCas9(D10A,H840A)融合evoCDA1脱氨酶的蛋白表达框,evoCDA1脱氨酶连接在dCas9(D10A,H840A)的N端,dCas9(D10A,H840A)的C端连接一个UGI,以T3T terminator为终止子。(2)esgRNA由大肠杆菌人工启动子pJ23119启动,以rrnB T2 terminator终止。(3)sgRNA靶标序列来自A. tumefaciens EHA105的recA基因(5’-TCCCAGATCGAACGGTCGTT-3’)。在远离PAM端的4 bp处有1个CAG密码子(Q26)作为CBE碱基编辑器C-T的测试靶点。
携带GUS报告基因的载体是本实验室保存的pCAMBIA3301基础载体,该载体p3301-GUS使用了CaMV35S启动子启动GUS蛋白表达,以NOS终止子终止。携带CRC报告基因的p3301-CRC载体是以实验室保存的pCAMBIA3301载体为基础构建的,使用NOS启动子启动ZmC1-ZmR1-ZmC1融合蛋白的表达,以ter-CRC终止子终止。
载体的构建采用同源重组的方法,同源重组引物设计的原则为,相邻2个连接片段的上游引物与下游引物之间至少有20 bp的同源序列,引物序列由生工生物工程(上海)股份有限公司合成。各个元件设计同源重组的引物后使用Phanta SE Super- Fidelity DNA Polymerase进行PCR扩增,琼脂糖凝胶回收片段后使用同源重组酶NEBuilder® HiFi DNA Assembly Master Mix进行连接,连接产物在大肠杆菌Trans-T1菌株感受态中转化。在37 ℃培养箱培养1 d后,挑取单克隆设计检测引物进行PCR扩增,并进行Sanger测序,以验证片段与载体是否连接成功。
1.2.2 农杆菌菌株感受态制备
将农杆菌接种于含适当抗生素的YEB固体培养基上,于28 °C培养2 d。挑取单菌落,接种于5 mL含抗生素的YEB液体培养基中,28 °C振荡培养过夜;取过夜培养菌液2 mL,接种于200 mL YEB培养基中,28 ℃,180转/min振荡培养至OD600=0.5;收集菌液5 000转/min离心5 min,4 ℃离心;加入10 mL 0.15 mol/L预冷NaCl悬浮农杆菌,5000转/min离心5 min,4 ℃离心;加入1 mL预冷的20.00 mmol/L CaCl2(10%甘油),悬浮细胞,于24 h内使用,或分装成每管50 μL,液氮速冻1 min后,置于-80 ℃保存备用。
1.2.3 农杆菌转化
按照Beef Extract(5 g/L)、Yeast Extract(1 g/L)、Peptone(5 g/L),MgSO4·7H2O(4 g/L)和Agar(12 g/L)的比例配制YEB培养基,之后加入利福平及筛选转化载体对应的抗生素。取50 μL农杆菌感受态细胞,冰上融化,加入1 μg质粒DNA,混匀。液氮速冻5 min,37 ℃热激5 min,立即置于冰上2 min。加入1 mL YEB液体培养基,28 ℃(180转/min)振荡培养2~3 h。取出100~200 μL菌液涂布于含适当抗生素的YEB固体培养基上,28 °C培养2~3 d。
1.2.4 农杆菌中碱基编辑载体消除
挑取EHA105(ΔrecA)突变体菌株的正确克隆,使用利福平/卡那霉素双抗性液体培养基振荡培养20 h至OD600= 0.6。取500 μL菌液加入50 mL利福平抗性的液体YEB培养基,连续转接培养10次后,取转接培养第10次的菌液稀释103倍,取50 μL涂布于利福平抗性固体培养基中。挑取同一单克隆分别点在利福平抗性、利福平/卡那霉素双抗性固体培养基中,培养2 d后,筛选到某一个单克隆在利福平抗性培养基中正常生长,而在利福平/卡那霉素双抗性固体培养基中没有生长的即为消除碱基编辑载体的目标菌株。
1.2.5 EHA105(ΔrecA)突变型菌株的DNA损伤测试
以培养完成后的EHA105和EHA105(ΔrecA)突变株2种菌液(OD600=0.6)为基础,每一个梯度稀释都在上一个梯度的基础上稀释10倍,对每一个梯度的菌液使用10 μL移液器取5 μL轻轻转移至分别含有0.005%烷化剂甲基磺酸甲酯(MMS)和无MMS的YEB培养基上,28 ℃培养2 d后采集照片。
1.2.6 农杆菌侵染玉米幼胚瞬时转化
农杆菌培养:将农杆菌接种于含适当抗生素的YEB固体培养基上,28 °C培养2 d。挑取单菌落,接种于5 mL含抗生素的YEB液体培养基中,28 °C振荡培养过夜。将过夜培养物按1:100比例接种于50 mL含抗生素的YEB液体培养基中,28 °C振荡培养至OD600=0.6~0.8。加入乙酰丁香酮(AS,200 μmol/L),继续培养4~6 h。5 000转/min离心10 min,收集菌体,用侵染培养基(AS,200 μmol/L)重悬至OD600= 0.5~1.0。
农杆菌侵染:将玉米幼胚浸泡在70%乙醇中1 min,转移至10% NaClO溶液中浸泡10~15 min,用无菌水冲洗3~5次,彻底去除消毒剂。在超净工作台中,用镊子和解剖刀从籽粒中分离出幼胚。将分离的幼胚浸入农杆菌悬浮液中,轻轻振荡10~30 min,用无菌滤纸吸干幼胚表面多余的菌液。将侵染后的幼胚置于共培养基上,22~25 ℃暗培养2~9 d后用含抗生素的无菌水清洗幼胚,去除表面农杆菌。
GUS染色:将幼胚浸入含0.3%甲醛的磷酸缓冲液(pH 7.0)中,室温固定30 min后将幼胚浸入GUS染色液中,37 ℃避光孵育数小时至过夜。用70%乙醇脱色,直至背景清晰。
1.2.7 碱基编辑农杆菌基因型鉴定
在其recA基因上目标靶位点的前后200~300 bp,设计PCR扩增引物序列(表1),扩增大小为517 bp,PCR扩增后产物进行Sanger测序。
表1 相关引物序列
Table 1
| 引物名称Primer name | 引物序列(5’-3’)Primer sequence (5’-3’) |
|---|---|
| Q26-HF1 | CCTAGGTATAATACTAGTTCCCAGATCGAACGGTCGTTGTTTCAGAGCTATGCTGGAAA |
| Q26-HF2 | TCCTCTAGAGTCGACCTGCAGGTTGACAGCTAGCTCAGTCCTAGGTATAATACTAGTTC |
| Q26-HF3 | TTCGAGCTCGGTACCCGGGGATCCTCTAGAGTCGACCTGCAGG |
| ScaffT2T-HR | TTAAAAAAAGGCCATCCGTCAGGATGGCCTTCTGCACCGACTCGGTGCCACTTTTTCA |
| Lac-flag-HF1 | CTTTATGCTTCCGGCTCGTATGTTAGGAGGTCTTTATCATGGACTATAAGGACCACGAC |
| Lac-flag-HF2 | CCATCCTGACGGATGGCCTTTTTTTAAGCTTTTTACACTTTATGCTTCCGGCTCGTATG |
| Flag-CDA1-HR | GAATCCGCACGTACTCGGCGTCGGTCTTATCGTCATCGTCTTTGTAA |
| Flag-CDA1-HF | GATGACGATAAGACCGACGCCGAGTACGTGCGGATTCAC |
| CDA1-cas9-HR | GAGGCCGATGGAGTACTTCTTGTCCACGGCGGGAGACTTTGTGGTGTGC |
| Cas9-HF | GACAAGAAGTACTCCATCGGCCTCGCCAT |
| UGI-T3T-HR1 | GAAAGAAGAAAGGGGCAATAACGCCCCACATTTTATACGAGCATCTTAATCTTGTTCTC |
| T3T-MaubI-HR2 | ATTCCCGATCTAGTAACATAGATGACACCGCGCGCGAGAAAGAAGAAAGGGGCAATAAC |
| T2-virB-HF | CCATCCTGACGGATGGCCTTTTTTTAAGCTTCGAGCATTTTCGTTCTGAGCCTGACCTG |
| VirB-flag-HR | GTCGTGGTCCTTATAGTCCATACCTTATCTCCTTAGCTCGCAACTA |
| VirB-flag-HF | AGTTGCGAGCTAAGGAGATAAGGTATGGACTATAAGGACCACGA |
| CDA1-cas9-HR | GAGGCCGATGGAGTACTTCTTGTCCACGGCGGGAGACTTTGTGGTGTGC |
| Lac-flag-HF1 | CTTTATGCTTCCGGCTCGTATGTTAGGAGGTCTTTATCATGGACTATAAGGACCACGAC |
| Lac-flag-HF2 | CCATCCTGACGGATGGCCTTTTTTTAAGCTTTTTACACTTTATGCTTCCGGCTCGTATG |
| Flag-T8e-HR | ACTCGTGGGAAAACTCCACCTCAGACTTATCGTCATCGTCTTTGTAA |
| TadA8e-HF | ACTCGTGGGAAAACTCCACCTCAGACTTATCGTCATCGTCTTTGTAA |
| Cas9-T3T-HR | AAAGAAGAAAGGGGCAATAACGCCCCACATTTTATACGTCGCCGCCGAGCTGGGAGAGG |
| TadA8e-HF | TCTGAGGTGGAGTTTTCCCACGAGT |
| TadA-cas9-HR | GAGGCCGATGGAGTACTTCTTGTCGTCTGTGGAGCTCTGGGCCTTCTTCTGA |
| JCF-3301 | CACTCATTAGGCACCCCAGG |
| JCR-Cas9 | AAAATCTCCTGGAGGTAGCAG |
| JCF-H840A | AAGAAGGGCATCCTCCAGAC |
| JCR-3301 | CTTTGATCCCGAGGGGAACC |
| JCF-Q26 | CAAATCCACACCGAGCTTGC |
| JCR-Q26 | GGCACGGAAAAACGAAAGCA |
2 结果与分析
2.1 不同ABE碱基编辑器在农杆菌EHA105精准编辑活性比较
表2 不同腺嘌呤脱氨酶变体
Table 2
| 变体 Variant | 位点Site | ||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 23 | 26 | 27 | 28 | 36 | 47 | 48 | 51 | 61 | 73 | 76 | 82 | 84 | 96 | 106 | 108 | 109 | 110 | 111 | |||||||||||||||||
| TadAWT | W | R | E | V | H | R | P | R | M | Y | I | V | L | H | A | D | A | K | T | ||||||||||||||||
| TadA8e | R | L | A | L | F | W | N | ||||||||||||||||||||||||||||
| TadA8.20 | R | L | A | L | Y | S | F | W | N | ||||||||||||||||||||||||||
| TadA8r | R | L | K | A | L | Y | S | W | G | R | H | ||||||||||||||||||||||||
| 变体 Variant | 位点Site | ||||||||||||||||||||||||||||||||||
| 114 | 119 | 122 | 123 | 126 | 127 | 146 | 147 | 149 | 152 | 154 | 155 | 156 | 157 | 158 | 161 | 166 | 167 | ||||||||||||||||||
| TadAWT | A | D | H | H | M | N | S | D | F | R | Q | E | I | K | A | K | T | D | |||||||||||||||||
| TadA8e | Y | C | Y | P | V | F | N | ||||||||||||||||||||||||||||
| TadA8.20 | C | R | P | R | V | F | N | ||||||||||||||||||||||||||||
| TadA8r | V | N | Y | I | K | C | R | P | R | V | F | ||||||||||||||||||||||||
图1
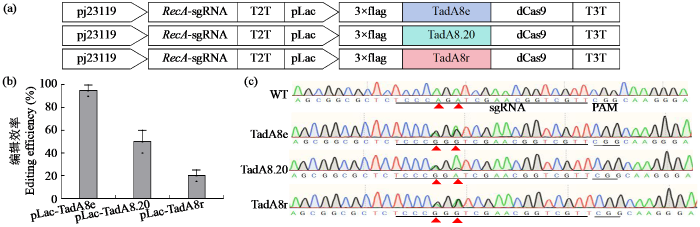
图1
不同ABE碱基编辑器介导的农杆菌EHA105精准编辑
Fig.1
Precise editing of Agrobacterium EHA105 mediated by different ABE base editors
为评估不同ABE碱基编辑器的编辑效率,将3种ABE系统分别转化农杆菌EHA105后,随机筛选20个单克隆进行靶位点PCR扩增及Sanger测序分析。基因分型结果(图1b)显示,TadA8e-dCas9系统编辑效率达95%(19/20),显著高于TadA8.20- dCas9(50%,10/20)和TadA8r-dCas9(20%,4/20)。值得注意的是,所有有效编辑均集中在靶序列的A5和A7位点(A→G转换),而在A11和A12位点未检测到编辑活性,这可能与脱氨酶的空间构象偏好或局部DNA二级结构特征相关。靶位点编辑Sanger测序结果(图1c)显示,所有突变基因型均为杂合或嵌合状态,未检测到纯合突变克隆。
2.2 CBE碱基编辑器介导的农杆菌EHA105 recA基因精准敲除
RecA作为细菌DNA修复与同源重组过程中的核心调控蛋白,在维持基因组稳定性中发挥关键作用,敲除recA基因可通过降低外源质粒在宿主细胞内的重组频率,提升外源质粒的表达稳定性。基于CBE碱基编辑系统,针对农杆菌EHA105菌株recA基因设计精准敲除载体,通过C→T碱基转换在Q26位点(CAG密码子)引入提前终止密码子(TAG)。靶位点的选择综合考虑了CBE系统的最适编辑窗口及突变位点对蛋白功能的影响,最终确定靶向recA基因N端功能域的20 bp序列(图2a)。研究构建了2种调控模式的CBE系统,将evoCDA1胞嘧啶脱氨酶变体[20]与dCas9蛋白N端融合,并在C端连接尿嘧啶糖基化酶抑制剂(UGI),分别由细菌组成型启动子pLac和农杆菌诱导型启动子pvirB驱动表达。sgRNA表达单元采用大肠杆菌人工启动子pJ23119进行组成型调控(图2b)。
图2
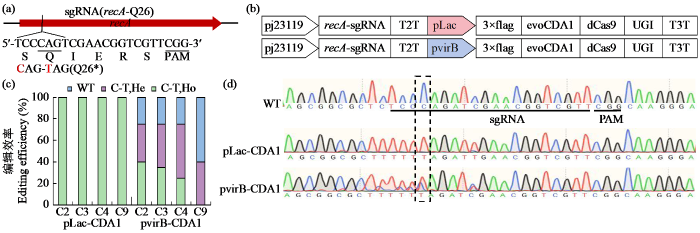
图2
CBE碱基编辑器介导的农杆菌EHA105 recA基因精准敲除
Fig.2
CBE base editor mediates precise knockout of Agrobacterium EHA105 recA gene
将pLac与pvirB驱动evoCDA1-dCas9-UGI的CBE碱基编辑载体分别转入EHA105菌株,随机筛选20个单克隆进行recA靶位点PCR扩增及Sanger测序分析。测序结果(图2c)显示,基于pLac启动子表达evoCDA1-dCas9-UGI的CBE碱基编辑器的编辑效率为100%(20/20),而pvirB启动子表达evoCDA1-dCas9-UGI的CBE碱基编辑器的编辑效率为75%(15/20)。峰图解析结果(图2d)表明,pLac启动子的CBE编辑器突变子均为纯合突变(100%),C→T编辑不仅发生在Q26编码密码子的C4位点(100%),还发生在靶点内的C2和C3位点(效率均为100%)。pvirB启动子的CBE编辑器突变子在目标C4位点纯合突变效率为25%(5/20),杂合突变效率为50%(10/20)。C→T编辑也发生在靶点内的C2、C3和C9位点,编辑效率相对较低,C2和C3位点处为75%(15/20),C9位点处为40%(8/20)。值得注意的是,2组系统均检测到靶点上游C-1位点的编辑。
2.3 EHA105(ΔrecA)突变型菌株对DNA损伤的敏感性测试
图3
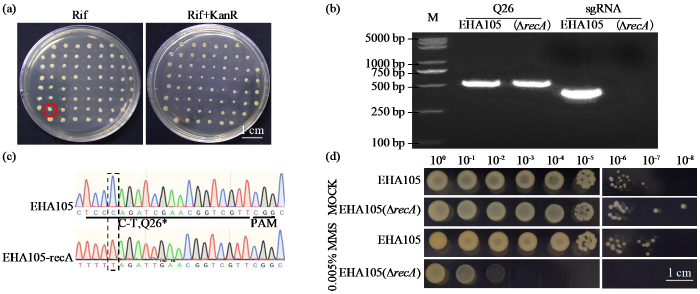
图3
EHA105(ΔrecA)突变型菌株的DNA损伤测试
(a) 卡那霉素敏感性测试;(b) 载体特异性引物扩增PCR;(c) recA突变位点Sanger测序验证;(d) DNA损伤(0.005%MMS)测试。
Fig.3
DNA damage testing of EHA105(ΔrecA) mutant strains
(a) Kanamycin sensitivity recovery test (b) PCR amplification of specific primers after eliminating the editing vector (c) Sanger sequencing verification of recA mutation sites (d) DNA damage (0.005% MMS) test.
为评估recA基因功能缺失对DNA损伤应答的影响,研究比较了野生型EHA105与EHA105(ΔrecA)突变株对烷化剂甲基磺酸甲酯(MMS)的敏感性。将2种菌株培养至对数生长期(OD600= 0.6),进行10倍梯度稀释后点样于含0.005% MMS的YEB平板,同步接种于无MMS的YEB平板作为对照组。结果表明,无MMS处理条件下,EHA105(ΔrecA)与野生型菌株生长速率及菌落形态无显著差异。在0.005% MMS处理条件下,野生型菌株的存活率同未处理组,而EHA105(ΔrecA)存活率骤降(图3d)。因此,农杆菌recA基因缺失导致菌株对DNA损伤高度敏感。
2.4 基于EHA105(ΔrecA)菌株的三元载体系统对玉米幼胚的瞬时侵染效率分析
基于recA基因敲除的农杆菌EHA105(ΔrecA)菌株,研究构建了三元载体系统。该系统由Ti质粒pTi542DT-DNA(含完整VIR基因簇,但缺失T-DNA转移能力)、辅助质粒(壮观霉素抗性)和T-DNA双元载体(卡那霉素抗性)3部分组成。辅助质粒携带pTi542DT-DNA来源VIR基因簇的复制子载体,分别采用pVS1和pRiA4复制子系统(图4a)。T-DNA双元载体分别包含载体p3301-GUS和p3301-CRC的报告基因(图4b)。其中,GUS基因编码β-葡萄糖醛酸乙酰转移酶,能够将5-溴-4-氯-3-吲哚-β-D-葡萄糖苷酸酯(X-Gluc)水解,生成蓝色的产物。CRC为ZmC1和ZmR1蛋白的嵌合体蛋白,可以激活花青素的生物合成。
图4
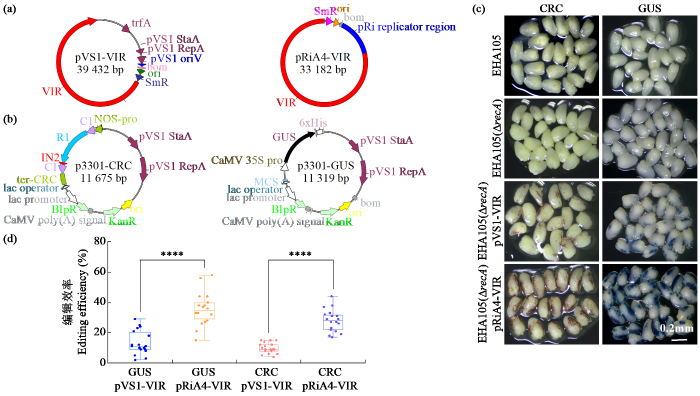
图4
基于EHA105(ΔrecA)菌株的三元载体系统对玉米幼胚的瞬时侵染
(a) pVS1或pRiA4复制子辅助质粒载体结构示意图;(b) 表达报告基因的双元载体p3301-GUS和p3301-CRC的结构示意图;(c) 基于EHA105(ΔrecA)菌株的三元载体系统对玉米幼胚的瞬时侵染;(d) 基于EHA105(ΔrecA)菌株的三元系统侵染效率(n=18);“****”表示差异极显著(P < 0.0001)。
Fig.4
Transient infection of maize immature embryos using the ternary vector system based on the EHA105(ΔrecA) strain
(a) schematic diagram of helper plasmid vector structures; (b) schematic diagram of reporter binary vectors; (c) transient infection of maize immature embryos using the ternary vector system based on the EHA105(ΔrecA) strain; (d) ternary systemic infection efficiency based on EHA105(ΔrecA) strain (n=18),“****”indicates extremely significant differences (P < 0.0001).
将2种辅助侵染质粒分别导入到EHA105和EHA105(ΔrecA)菌株中,进一步在EHA105、携带或不携带辅助侵染质粒的EHA105(ΔrecA)农杆菌中分别导入p3301-GUS和p3301-CRC,形成三元表达载体。利用以上农杆菌瞬时侵染授粉后12 d的玉米幼胚,共培养3 d后观察报告基因的表达。以单个胚染色面积的占比来统计侵染效率,利用Photoshop的像素数计算出图像中的染色面积的占比[21]。研究结果(图4c、d)显示,未导入辅助侵染质粒组,recA基因敲除没有降低农杆菌的侵染能力。导入辅助质粒后可以显著增强农杆菌的侵染能力,其中pRiA4-VIR辅助侵染质粒介导的农杆菌的的侵染效率为28%~35%,显著高于pVS1-VIR(10%~13%)。
3 讨论
农杆菌介导的遗传转化是禾本科植物基因改造的重要工具,其推动的基因编辑技术已在农作物育种领域引发革命性突破。然而,尽管农杆菌广泛应用于植物分子生物学研究,但现有菌株资源与技术协同开发尚未充分适配重要作物的转化需求。本研究通过优化ABE和CBE碱基编辑系统,成功实现了农杆菌EHA105基因组的高效精准编辑,并基于recA基因敲除菌株构建了新型三元载体系统。
本研究发现,ABE系统中TadA8e-dCas9的编辑效率(95%)显著高于TadA8.20(50%)和TadA8r(20%),且其性能趋势与动植物细胞的报道[17,19]一致。这表明农杆菌凭借其短生命周期、单倍体基因组特性和成熟的遗传操作体系,可作为快速筛选碱基编辑器的理想模型。此外,启动子类型显著影响碱基系统的编辑效率和突变纯合度。基于组成型pLac启动子的CBE系统编辑效率达100%,且突变均为纯合型,而诱导型pvirB系统效率仅为75%,且仅产生杂合/嵌合突变。这一差异源于两者的表达动力学特性,pLac的持续高表达可覆盖DNA复制全程,确保脱氨酶稳定作用于所有染色体拷贝,从而实现同步编辑;而pvirB依赖VirA/VirG信号系统激活[22],其表达的波动导致脱氨酶可能仅在部分细胞周期中有效,最终引发异质性编辑,表明应根据需要选择合适的脱氨酶变体及启动子。
EHA105(ΔrecA)菌株虽成功消除了外源质粒的重组风险,但其对MMS的高度敏感性揭示了recA缺失的潜在局限性。RecA作为同源重组修复的核心蛋白,其功能缺失导致菌株对DNA损伤剂耐受性骤降,可能限制其在含基因毒性成分的植物组织中的应用。然而,在常规转化体系中,recA缺失通过稳定辅助质粒显著提升了侵染效率,且未影响基础生长速率。因此,在实际应用中需根据转化环境权衡recA编辑的利弊,必要时可通过添加DNA保护剂或优化共培养条件以弥补修复缺陷带来的负面影响。
基于EHA105(ΔrecA)构建的三元载体系统,通过分离VIR基因簇(辅助质粒)与T-DNA(双元载体),提升了侵染效率。该过程中需要协调辅助质粒与T-DNA质粒的兼容性,试验采用的双元表达T-DNA质粒同样属于pVS1复制子系统,携带pRiA4复制子的辅助质粒在侵染效率上优于pVS1复制子系统。本研究成功开发了农杆菌基因组精准编辑技术体系,实现了对农杆菌关键功能基因的高效定向改造,为农杆菌工程化改良及植物遗传转化效率提升提供了创新性解决方案。未来可进一步探索优化表达策略以协调辅助质粒和T-DNA质粒表达稳定性。
4 结论
系统比较3种ABE变体在农杆菌中的编辑效率,发现其效率差异与动植物细胞中的报道趋势高度一致,表明农杆菌可作为快速筛选和优化新型碱基编辑器的标准化测试平台。通过CBE碱基编辑器在农杆菌的recA基因上的引入提前终止突变,获得recA功能缺失突变的EHA105菌株,突变效率达100%。在EHA105(ΔrecA)农杆菌菌株基础上引入辅助侵染质粒与报告基因系统,显著提升了农杆菌介导的玉米幼胚的瞬时侵染效率。
参考文献
Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana
DOI:10.1046/j.1365-313x.1998.00343.x
URL
[本文引用: 1]

The\n \n Agrobacterium\n \n vacuum infiltration method has made it possible to transform\n \n Arabidopsis thaliana\n \n without plant tissue culture or regeneration. In the present study, this method was evaluated and a substantially modified transformation method was developed. The labor‐intensive vacuum infiltration process was eliminated in favor of simple dipping of developing floral tissues into a solution containing\n \n Agrobacterium tumefaciens\n \n, 5% sucrose and 500 microliters per litre of surfactant Silwet L‐77. Sucrose and surfactant were critical to the success of the floral dip method. Plants inoculated when numerous immature floral buds and few siliques were present produced transformed progeny at the highest rate. Plant tissue culture media, the hormone benzylamino purine and pH adjustment were unnecessary, and\n \n Agrobacterium\n \n could be applied to plants at a range of cell densities. Repeated application of\n \n Agrobacterium\n \n improved transformation rates and overall yield of transformants approximately twofold. Covering plants for 1 day to retain humidity after inoculation also raised transformation rates twofold. Multiple ecotypes were transformable by this method. The modified method should facilitate high‐throughput transformation of\n \n Arabidopsis\n \n for efforts such as T‐DNA gene tagging, positional cloning, or attempts at targeted gene replacement.
Agrobacterium -mediated transformation of rice using immature embryos or calli induced from mature seed
DOI:10.1038/nprot.2008.46
PMID:18451790
Here, we provide comprehensive, highly efficient protocols for Agrobacterium tumefaciens-mediated transformation of a wide range of rice genotypes. Methods that use either immature embryos (japonica and indica rice) or calli (japonica cultivars and the indica cultivar, Kasalath) as a starting material for inoculation with Agrobacterium are described. Immature embryos are pretreated with heat and centrifugal force, which significantly enhances the efficiency of gene transfer, and then infected with Agrobacterium. Callus is induced from mature seeds and infected. Transformed cells proliferated from these tissues are selected on the basis of hygromycin resistance, and transgenic plants are eventually regenerated. A single immature japonica or Kasalath embryo will produce between 10 and 18 independent transgenic plants; for other non-Kasalath indica varieties, the number of transgenic plants expected will be between 5 and 13. For japonica and Kasalath, transformants should be obtained from between 50 and 90% of calli. From inoculation with Agrobacterium to transplanting to soil will take 55 d for japonica and Kasalath, and 74 d for indica other than Kasalath using the immature embryo method, and 50 d for japonica and Kasalath using the callus method.
Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew
DOI:10.1038/nbt.2969
PMID:25038773
Sequence-specific nucleases have been applied to engineer targeted modifications in polyploid genomes, but simultaneous modification of multiple homoeoalleles has not been reported. Here we use transcription activator-like effector nuclease (TALEN) and clustered, regularly interspaced, short palindromic repeats (CRISPR)-Cas9 (refs. 4,5) technologies in hexaploid bread wheat to introduce targeted mutations in the three homoeoalleles that encode MILDEW-RESISTANCE LOCUS (MLO) proteins. Genetic redundancy has prevented evaluation of whether mutation of all three MLO alleles in bread wheat might confer resistance to powdery mildew, a trait not found in natural populations. We show that TALEN-induced mutation of all three TaMLO homoeologs in the same plant confers heritable broad-spectrum resistance to powdery mildew. We further use CRISPR-Cas9 technology to generate transgenic wheat plants that carry mutations in the TaMLO-A1 allele. We also demonstrate the feasibility of engineering targeted DNA insertion in bread wheat through nonhomologous end joining of the double-strand breaks caused by TALENs. Our findings provide a methodological framework to improve polyploid crops.
Agrobacterium-mediated transformation of maize
DOI:10.1038/nprot.2007.241
PMID:17585302
Maize may be transformed very efficiently using Agrobacterium tumefaciens-mediated methods. The most critical factor in the transformation protocol is the co-cultivation of healthy immature embryos of the correct developmental stage with A. tumefaciens; the embryos should be collected only from vigorous plants grown in well-conditioned glasshouses. With the protocol described here, approximately 50% of immature embryos from the inbred line A188 and 15% from inbred lines A634, H99 and W117 will produce transformants. About half of the transformed plants are expected to carry one or two copies of the transgenes, which are inherited by the progeny in a mendelian fashion. More than 90% of transformants are expected to be normal in morphology. The protocol takes about 3 months from the start of co-cultivation to the planting of transformants into pots.
Targeted mutagenesis in soybean using the CRISPR-Cas 9 system
DOI:10.1038/srep10342
[本文引用: 1]
Genome editing is a valuable technique for gene function analysis and crop improvement. Over the past two years, the CRISPR-Cas9 system has emerged as a powerful tool for precisely targeted gene editing. In this study, we predicted 11 U6 genes in soybean (Glycine max L.). We then constructed two vectors (pCas9-GmU6-sgRNA and pCas9-AtU6-sgRNA) using the soybean U6-10 and Arabidopsis U6-26 promoters, respectively, to produce synthetic guide RNAs (sgRNAs) for targeted gene mutagenesis. Three genes, Glyma06g14180, Glyma08g02290 and Glyma12g37050, were selected as targets. Mutations of these three genes were detected in soybean protoplasts. The vectors were then transformed into soybean hairy roots by Agrobacterium rhizogenes infection, resulting in efficient target gene editing. Mutation efficiencies ranged from 3.2–9.7% using the pCas9-AtU6-sgRNA vector and 14.7–20.2% with the pCas9-GmU6-sgRNA vector. Biallelic mutations in Glyma06g14180 and Glyma08g02290 were detected in transgenic hairy roots. Off-target activities associated with Glyma06g14180 and Glyma12g37050 were also detected. Off-target activity would improve mutation efficiency for the construction of a saturated gene mutation library in soybean. Targeted mutagenesis using the CRISPR-Cas9 system should advance soybean functional genomic research, especially that of genes involved in the roots and nodules.
Agrobacterium -mediated genetic transformation of plants: biology and biotechnology
DOI:10.1016/j.copbio.2006.01.009
PMID:16459071
[本文引用: 1]
Agrobacterium-mediated genetic transformation is the dominant technology used for the production of genetically modified transgenic plants. Extensive research aimed at understanding and improving the molecular machinery of Agrobacterium responsible for the generation and transport of the bacterial DNA into the host cell has resulted in the establishment of many recombinant Agrobacterium strains, plasmids and technologies currently used for the successful transformation of numerous plant species. Unlike the role of bacterial proteins, the role of host factors in the transformation process has remained obscure for nearly a century of Agrobacterium research, and only recently have we begun to understand how Agrobacterium hijacks host factors and cellular processes during the transformation process. The identification of such factors and studies of these processes hold great promise for the future of plant biotechnology and plant genetic engineering, as they might help in the development of conceptually new techniques and approaches needed today to expand the host range of Agrobacterium and to control the transformation process and its outcome during the production of transgenic plants.
Agrobacterium tumefaciens gene transfer: how a plant pathogen hacks the nuclei of plant and nonplant organisms
DOI:10.1094/PHYTO-12-14-0380-RVW
PMID:26151736
[本文引用: 1]
Agrobacterium species are soilborne gram-negative bacteria exhibiting predominantly a saprophytic lifestyle. Only a few of these species are capable of parasitic growth on plants, causing either hairy root or crown gall diseases. The core of the infection strategy of pathogenic Agrobacteria is a genetic transformation of the host cell, via stable integration into the host genome of a DNA fragment called T-DNA. This genetic transformation results in oncogenic reprogramming of the host to the benefit of the pathogen. This unique ability of interkingdom DNA transfer was largely used as a tool for genetic engineering. Thus, the artificial host range of Agrobacterium is continuously expanding and includes plant and nonplant organisms. The increasing availability of genomic tools encouraged genome-wide surveys of T-DNA tagged libraries, and the pattern of T-DNA integration in eukaryotic genomes was studied. Therefore, data have been collected in numerous laboratories to attain a better understanding of T-DNA integration mechanisms and potential biases. This review focuses on the intranuclear mechanisms necessary for proper targeting and stable expression of Agrobacterium oncogenic T-DNA in the host cell. More specifically, the role of genome features and the putative involvement of host's transcriptional machinery in relation to the T-DNA integration and effects on gene expression are discussed. Also, the mechanisms underlying T-DNA integration into specific genome compartments is reviewed, and a theoretical model for T-DNA intranuclear targeting is presented.
Agrobacterium ‐mediated transformation of cereals: a promising approach crossing barriers
Cereal crops have been the primary targets for improvement by genetic transformation because of their worldwide importance for human consumption. For a long time, many of these important cereals were difficult to genetically engineer, mainly as a result of their inherent limitations associated with the resistance to Agrobacterium infection and their recalcitrance to in vitro regeneration. The delivery of foreign genes to rice plants via Agrobacterium tumefaciens has now become a routine technique. However, there are still serious handicaps with Agrobacterium-mediated transformation of other major cereals. In this paper, we review the pioneering efforts, existing problems and future prospects of Agrobacterium-mediated genetic transformation of major cereal crops, such as rice, maize, wheat, barley, sorghum and sugarcane.
The plant signal salicylic acid shuts down expression of the vir regulon and activates quormone- quenching genes in Agrobacterium
Identification of the signal molecules produced by wounded plant cells that activate T-DNA transfer in Agrobacterium tumefaciens
DOI:10.1038/318624a0 [本文引用: 1]
An improved ternary vector system for Agrobacterium-mediated rapid maize transformation
DOI:10.1007/s11103-018-0732-y [本文引用: 1]
A novel ternary vector system united with morphogenic genes enhances CRISPR/Cas delivery in maize
DOI:10.1104/pp.19.00767
PMID:31558579
[本文引用: 1]
The lack of efficient delivery methods is a major barrier to clustered regularly interspaced short palindromic repeats/CRISPR-associated protein (CRISPR/Cas)-mediated genome editing in many plant species. Combinations of morphogenic regulator (MR) genes and ternary vector systems are promising solutions to this problem. In this study, we first demonstrated that MR vectors greatly enhance maize () transformation. We then tested a CRISPR/Cas9 MR vector in maize and found that the MR and CRISPR/Cas9 modules have no negative influence on each other. Finally, we developed a novel ternary vector system to integrate the MR and CRISPR/Cas modules. Our ternary vector system is composed of new pGreen-like binary vectors, here named pGreen3, and a pVS1-based virulence helper plasmid, which also functions as a replication helper for the pGreen3 vectors in The pGreen3 vectors were derived from the plasmid pRK2 and display advantages over pGreen2 vectors regarding both compatibility and stability. We demonstrated that the union of our ternary vector system with MR gene modules has additive effects in enhancing maize transformation and that this enhancement is especially evident in the transformation of recalcitrant maize inbred lines. Collectively, our ternary vector system-based tools provide a user-friendly solution to the low efficiency of CRISPR/Cas delivery in maize and represent a basic platform for developing efficient delivery tools to use in other plant species recalcitrant to transformation.© 2019 American Society of Plant Biologists. All Rights Reserved.
Motoring along with the bacterial RecA protein
The recombinases of the RecA family are often viewed only as DNA-pairing proteins - they bind to one DNA segment, align it with homologous sequences in another DNA segment, promote an exchange of DNA strands and then dissociate. To a first approximation, this description seems to fit the eukaryotic (Rad51 and Dmc1) and archaeal (RadA) RecA homologues. However, the bacterial RecA protein does much more, coupling ATP hydrolysis with DNA-strand exchange in a manner that greatly expands its repertoire of activities. This article explores the protein activities and experimental results that have identified RecA as a motor protein.
Homologous recombination in Escherichia coli: dependence on substrate length and homology
DOI:10.1093/genetics/112.3.441
PMID:3007275
[本文引用: 1]
We studied the in vivo recombination between homologous DNA sequences cloned in phage lambda and a pBR322-derived plasmid by assaying for the formation of phage-plasmid cointegrates by a single (or an odd number of) reciprocal exchange. (1) Recombination proceeds by the RecBC pathway in wild-type cells and by low levels of a RecF-dependent pathway in recBC- cells. The RecE pathway appears not to generate phage-plasmid cointegrates. (2) Recombination is linearly dependent on the length of the homologous sequences. In both RecBC and RecF-dependent pathways there is a minimal length, called the minimal efficient processing segment (MEPS), below which recombination becomes inefficient. The length of MEPS is between 23-27 base pairs (bp) and between 44-90 bp for the RecBC- and RecF-dependent pathways, respectively. A model, based on overlapping MEPS, of the correlation of genetic length with physical length is presented. The bases for the different MEPS length of the two pathways are discussed in relationship to the enzymes specific to each pathway. (3) The RecBC and the RecF-dependent pathways are each very sensitive to substrate homology. In wild-type E. coli, reduction of homology from 100% to 90% decreases recombinant frequency over 40-fold. The homology dependence of the RecBC and RecF-dependent pathways are similar. This suggests that a component common to both, probably recA, is responsible for the recognition of homology.
Phage-assisted evolution of an adenine base editor with improved Cas domain compatibility and activity
DOI:10.1038/s41587-020-0453-z
PMID:32433547
[本文引用: 2]
Applications of adenine base editors (ABEs) have been constrained by the limited compatibility of the deoxyadenosine deaminase component with Cas homologs other than SpCas9. We evolved the deaminase component of ABE7.10 using phage-assisted non-continuous and continuous evolution (PANCE and PACE), which resulted in ABE8e. ABE8e contains eight additional mutations that increase activity (k) 590-fold compared with that of ABE7.10. ABE8e offers substantially improved editing efficiencies when paired with a variety of Cas9 or Cas12 homologs. ABE8e is more processive than ABE7.10, which could benefit screening, disruption of regulatory regions and multiplex base editing applications. A modest increase in Cas9-dependent and -independent DNA off-target editing, and in transcriptome-wide RNA off-target editing can be ameliorated by the introduction of an additional mutation in the TadA-8e domain. Finally, we show that ABE8e can efficiently install natural mutations that upregulate fetal hemoglobin expression in the BCL11A enhancer or in the the HBG promoter in human cells, targets that were poorly edited with ABE7.10. ABE8e augments the effectiveness and applicability of adenine base editing.
Directed evolution of adenine base editors with increased activity and therapeutic application
DOI:10.1038/s41587-020-0491-6
PMID:32284586
[本文引用: 1]
The foundational adenine base editors (for example, ABE7.10) enable programmable A•T to G•C point mutations but editing efficiencies can be low at challenging loci in primary human cells. Here we further evolve ABE7.10 using a library of adenosine deaminase variants to create ABE8s. At NGG protospacer adjacent motif (PAM) sites, ABE8s result in ~1.5× higher editing at protospacer positions A5-A7 and ~3.2× higher editing at positions A3-A4 and A8-A10 compared with ABE7.10. Non-NGG PAM variants have a ~4.2-fold overall higher on-target editing efficiency than ABE7.10. In human CD34 cells, ABE8 can recreate a natural allele at the promoter of the γ-globin genes HBG1 and HBG2 with up to 60% efficiency, causing persistence of fetal hemoglobin. In primary human T cells, ABE8s achieve 98-99% target modification, which is maintained when multiplexed across three loci. Delivered as messenger RNA, ABE8s induce no significant levels of single guide RNA (sgRNA)-independent off-target adenine deamination in genomic DNA and very low levels of adenine deamination in cellular mRNA.
An adenine base editor variant expands context compatibility
DOI:10.1038/s41587-023-01994-3 [本文引用: 2]
Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions
DOI:10.1038/nbt.3803
PMID:28191901
[本文引用: 1]
Base editing induces single-nucleotide changes in the DNA of living cells using a fusion protein containing a catalytically defective Streptococcus pyogenes Cas9, a cytidine deaminase, and an inhibitor of base excision repair. This genome editing approach has the advantage that it does not require formation of double-stranded DNA breaks or provision of a donor DNA template. Here we report the development of five C to T (or G to A) base editors that use natural and engineered Cas9 variants with different protospacer-adjacent motif (PAM) specificities to expand the number of sites that can be targeted by base editing 2.5-fold. Additionally, we engineered base editors containing mutated cytidine deaminase domains that narrow the width of the editing window from ∼5 nucleotides to as little as 1-2 nucleotides. We thereby enabled discrimination of neighboring C nucleotides, which would otherwise be edited with similar efficiency, and doubled the number of disease-associated target Cs able to be corrected preferentially over nearby non-target Cs.
virA and virG control the plant- induced activation of the T-DNA transfer process of A. tumefaciens
The Ti plasmid vir loci of Agrobacterium tumefaciens are transcriptionally activated in response to signal molecules produced by plant cells to initiate the T-DNA transfer process. We show that the pTiA6 vir loci are organized as a single regulon whose induction by plants is controlled by virA and virG. Mutations in virA result in attenuated induction. This locus is constitutively transcribed and noninducible. Mutations in virG eliminate vir induction. This locus is constitutively transcribed, plant-inducible, and self-regulated in a complex fashion, and it produces two distinct and differentially regulated transcripts. virA is proposed to encode a transport protein for the plant signal molecule, and virG a positive regulatory protein that together with the plant molecule activates vir expression.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}