
作物杂志,2024, 第4期: 121–129 doi: 10.16035/j.issn.1001-7283.2024.04.015
芝麻蒴果光合作用对粒重形成相关性状的贡献率分析
赵俊1( ), 段先琴2, 陈艳妮1, 汪文倩1, 曾袁琳珺1, 危文亮1(), 孙建3()
), 段先琴2, 陈艳妮1, 汪文倩1, 曾袁琳珺1, 危文亮1(), 孙建3()
- 1长江大学农学院,434025,湖北荆州
2武汉现代农业教育中心,430040,湖北武汉
3江西省农业科学院作物研究所,330200,江西南昌
Analysis of the Contribution Rate of Capsule Photosynthesis to the Traits Related to Grain Weight Formation in Sesamum indicum
Zhao Jun1(), Duan Xianqin2, Chen Yanni1, Wang Wenqian1, Zeng Yuanlinjun1, Wei Wenliang1(), Sun Jian3()
- 1College of Agriculture, Yangtze University, Jingzhou 434025, Hubei, China
2Wuhan Modern Agricultural Education Center, Wuhan 430040, Hubei, China
3Institute of Crop Research, Jiangxi Academy of Agricultural Sciences, Nanchang 330200, Jiangxi, China
摘要:
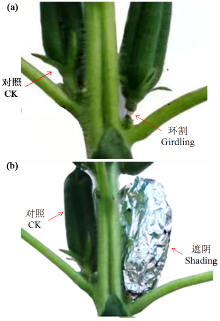
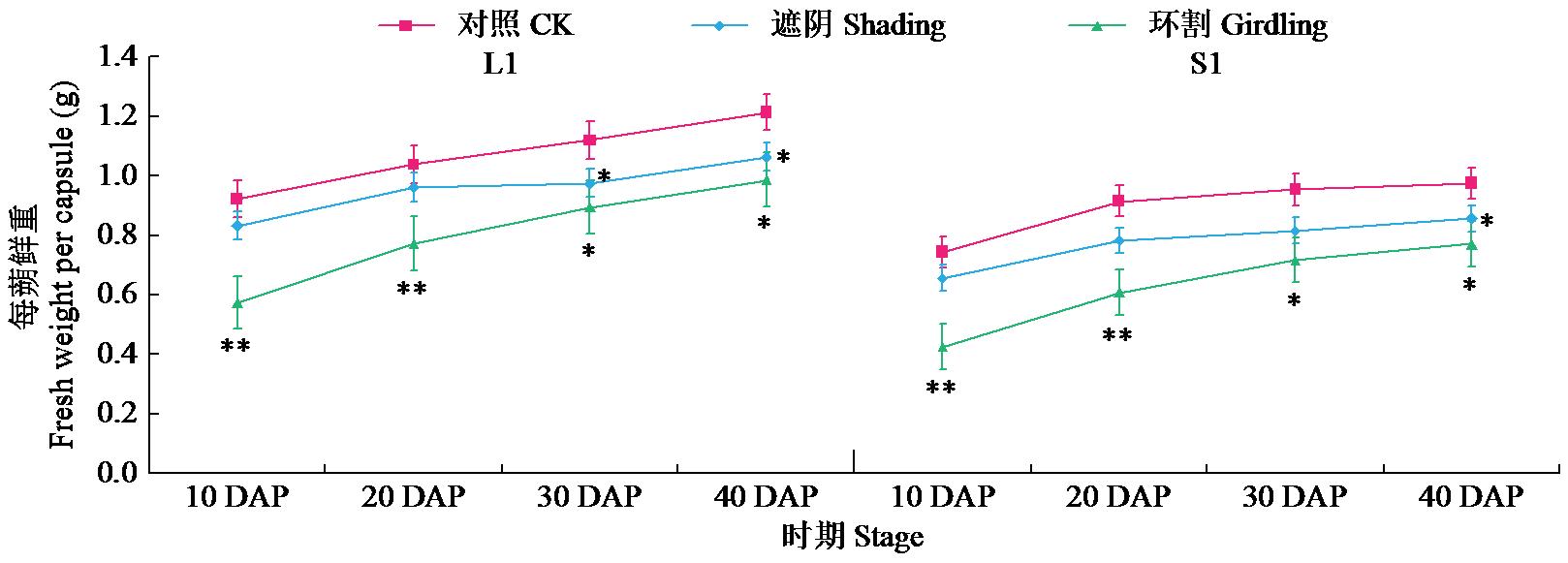
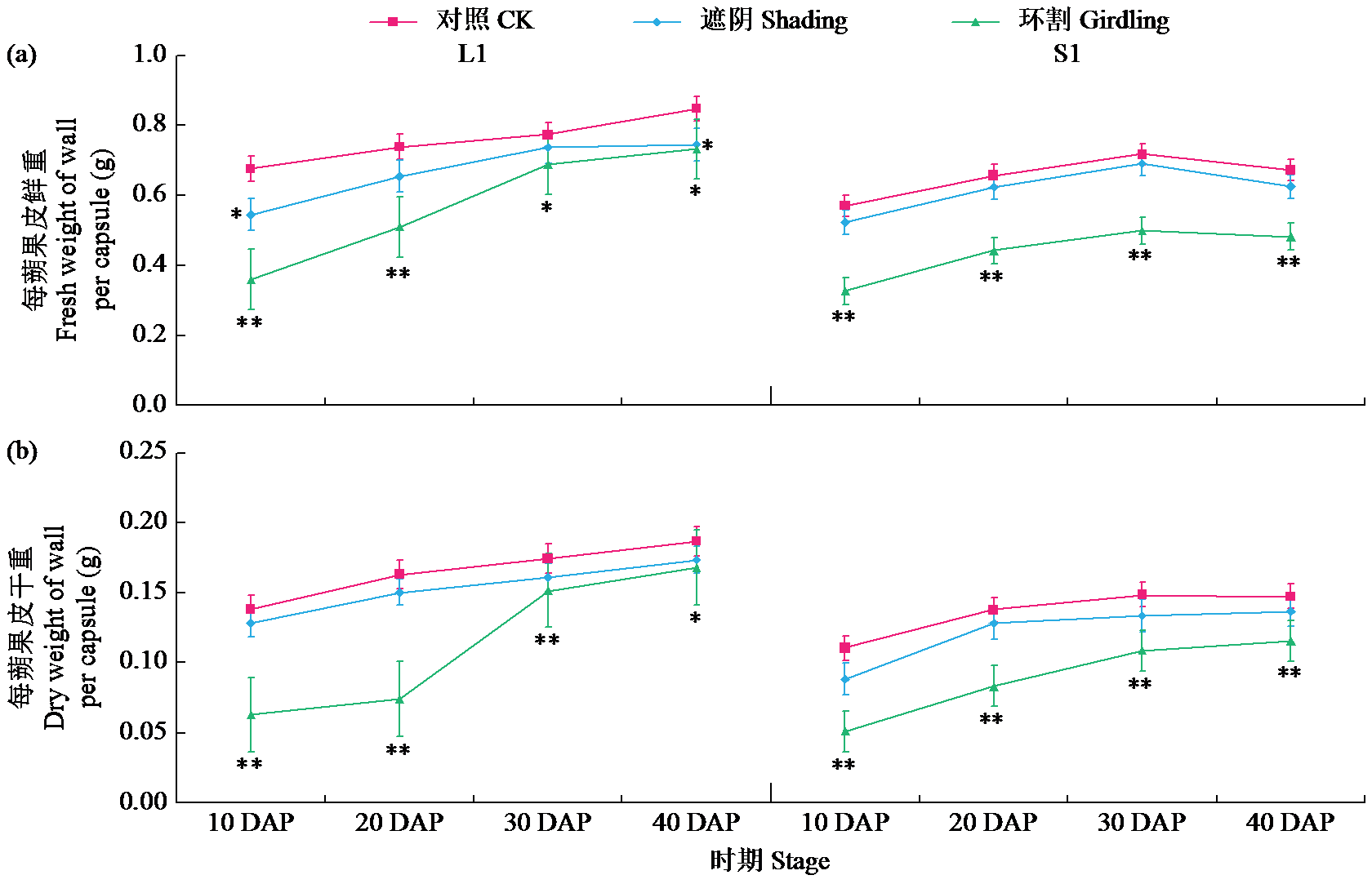
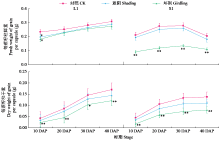
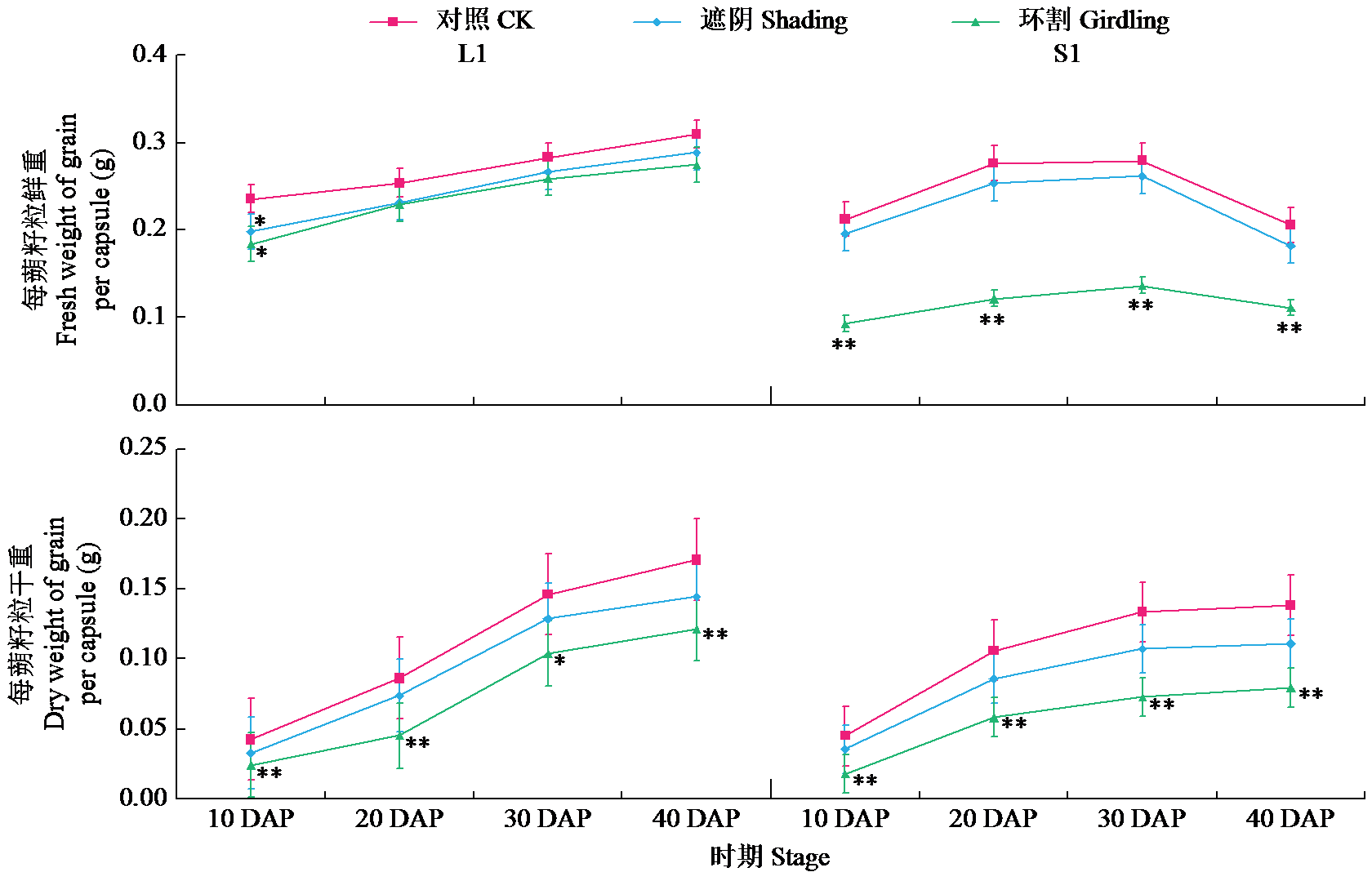
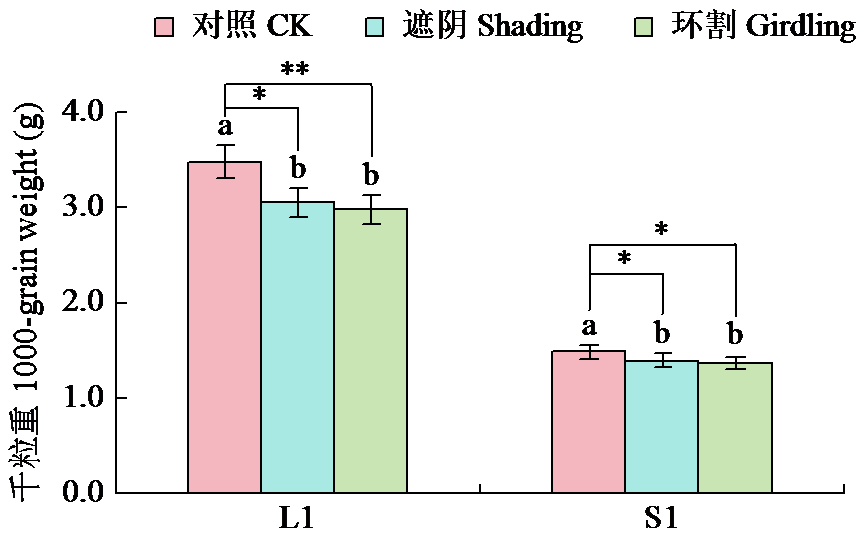
通过遮阴和环割处理分析了不同粒重芝麻材料的蒴果光合作用对每蒴鲜重、每蒴果皮鲜重和干重、每蒴籽粒鲜重和干重5个粒重形成相关性状的影响及贡献率大小。结果表明,遮阴处理下,除每蒴果皮鲜重外,其他4个性状与对照基本无显著差异,表明蒴果光合作用被抑制后能迅速被叶茎光合作用补偿;授粉后40 d(40 DAP)时大粒材料(L1)和小粒材料(S1)蒴果光合作用对每蒴果皮干重、每蒴籽粒干重的贡献率均较为接近,意味着不同粒重芝麻材料的叶茎光合作用对这些性状的贡献基本接近,且粒重差异的形成可能与蒴果的光合贡献有关。环割处理下,5个性状均与对照有显著差异,表明环割处理后叶茎光合作用不能完全被蒴果所补偿,且对蒴果发育和粒重形成有显著影响;而蒴果光合作用对每蒴鲜重、每蒴果皮干重、每蒴籽粒干重的贡献率都随着蒴果发育而呈增加趋势,意味着蒴果光合作用可能具有补偿增强效应;尤其在40 DAP时期,L1蒴果光合作用对每蒴果皮干重、每蒴籽粒干重的贡献率极显著高于S1,揭示大粒材料的粒重形成与其蒴果的较高光合贡献率有关。


| [1] | Burstin T J. Reserve accumulation in legume grains. Comptes Rendus Biologies, 2008, 331(10):755-762. |
| [2] | Aschan G, Pfanz H. Non-foliar photosynthesisa strategy of additional carbon acquisition. Flora, 2003, 198(2):81-97. |
| [3] | Mogensen V O, Jensen C R, Mortensen G, et al. Pod photosynthesis and drought adaptation of field grown rape (Brassica napus L.). European Journal of Agronomy, 1997, 6(3/ 4):295-307. |
| [4] | Hua W, Li R J, Zhan G M, et al. Maternal control of seed oil content in Brassica napus: the role of silique wall photosynthesis. The Plant Journal, 2011, 69(3):432-444. |
| [5] | 杨阳, 苍晶, 王学东, 等. 大豆豆荚光合特性及其对产量的贡献. 东北农业大学学报, 2008, 39(12):51-56. |
| [6] | Crookston R K, O'Toole J, Ozbun J L. Characterization of the bean pod as a photosynthetic organ. Crop Science, 1974, 14(5):708-712. |
| [7] |
Wang H, Hou L Y, Wang M Y, et al. Contribution of the pod wall to seed grain filling in alfalfa. Scientific Reports, 2016, 6:26586.
doi: 10.1038/srep26586 pmid: 27210048 |
| [8] | Zhang W X, Mao P S, Li Y, et al. Assessing of the contributions of pod photosynthesis to carbon acquisition of seed in alfalfa (Medicago sativa L.) open. Scientific Reports, 2019, 7:420-426. |
| [9] | Imaizumi N, Usuda H, Nakamoto H, et al. Changes in the rate of photosynthesis during grain filling and the enzymatic activities associated with the photosynthetic carbon metabolism in rice panicles. Plant & Cell Physiology, 1990, 31(6):835-844. |
| [10] |
Zhou B W, Serret M D, Elazab A, et al. Wheat ear carbon assimilation and nitrogen remobilization contribute significantly to grain yield. Journal of Integrative Plant Biology, 2016, 58 (11):914-926.
doi: 10.1111/jipb.12478 |
| [11] | Bort J, Brown R H, Araus J L. Refixation of respiratory CO2 in the ears of C3 cereals. Journal of Experimental Botany, 1996, 47 (10):1567-1575. |
| [12] |
Sui X L, Shan N, Hu L P, et al. The complex character of photosynthesis in cucumber fruit. Journal of Experimental Botany, 2017, 68(7):1625-1637.
doi: 10.1093/jxb/erx034 pmid: 28369547 |
| [13] | Piechulla B, Gruissem W. Diurnal mRNA fluctuations of nuclear and plastid genes in developing tomato fruits. The EMBO Journal, 1987, 6(12):3593-3599. |
| [14] | Hu Y Y, Zhang Y L, Luo H H, et al. Important photosynthetic contribution from the non-foliar green organs in cotton at the late growth stage. Planta, 2011, 235(2):325-336. |
| [15] | Wullschleger S D, Oosterhuis D M. Photosynthesis, transpiration, and water-use efficiency of cotton leaves and fruit. Photosynthetica, 1991, 25:505-515. |
| [16] | Zhang Y, Mulpuri S, Liu A Z, et al. Photosynthetic capacity of the capsule wall and its contribution to carbon fixation and seed yield in castor (Ricinus communis L.). Acta Physiologiae Plantarum, 2016, 38(10):1-12. |
| [17] | Kong L G, Wang F H, Feng B, et al. The structural and photosynthetic characteristics of the exposed peduncle of wheat (Triticum aestivum L.): an important photosynthate source for grain-filling. BMC Plant Biology, 2010, 10(1):141. |
| [18] | Wang Z M, Wei A L, Zheng D M. Photosynthetic characteristics of non-leaf organs of winter wheat cultivars differing in ear type and their relationship with grain mass per ear. Photosynthetica, 2001, 39(2):239-244. |
| [19] | Diepenbrock W. Yield analysis of winter oilseed rape (Brassica napus L.): a review. Field Crops Research, 2000, 67(1):35-49. |
| [20] |
Dubousset L, Etienne P, Avice J C. Is the remobilization of S and N reserves for seed filling of winter oilseed rape modulated by sulphate restrictions occurring at different growth stages?. Journal of Experimental Botany, 2010, 61(15):4313-4324.
doi: 10.1093/jxb/erq233 pmid: 20693411 |
| [21] |
Namiki M. Nutraceutical functions of sesame: a review. Critical Reviews in Food Science and Nutrition, 2007, 47(7):651-673.
pmid: 17943496 |
| [22] | Day J S. Development and maturation of sesame seeds and capsules. Field Crops Research, 2000, 67(1):1-9. |
| [23] | 陈艳妮, 徐鸿儒, 赵云燕, 等. 芝麻蒴果发育过程中蒴果皮的光合作用及籽粒充实的变化研究. 植物生理学报, 2022, 58 (4):723-732. |
| [24] | Duffus C M, Cochrane M P. Formation of the barley grain- morphology, physiology, and biochemistry//MacGregor A W,Bhatty R S (eds) Barley: Chemistry and Technology. American Association of Cereal Chemists,St. Paul, 1993:31-72. |
| [25] | Xu H L, Gauthier L, Gosselin Y D, et al. Photosynthesis in leaves, fruits, stem and petioles of greenhouse-grown tomato plants. Photosynthetica, 1997, 33(1):113-123. |
| [26] | Luo M, Dennis E S, Berger F, et al. MINISEED3 (MINI3), a WRKY family gene, and HAIKU2 (IKU2), a leucine-rich repeat (LRR) KINASE gene,are regulators of seed size in Arabidopsis. Proceedings of the National Academy of Sciences of the United States, 2005, 102 (48):17531-17536. |
| [27] | Martin A, Lee J, Kichey T, et al. Two cytosolic glutamine synthetase isoforms of maize are specifically involved in the control of grain production. The Plant Cell, 2006, 18(11):3252-3274. |
| [28] | Song X J, Huang W, Shi M, et al. A QTL for rice grain width and weight encodes a previously unknown RING-type E 3 ubiquitin ligase. Nature Genetics, 2007, 39(5):623-630. |
| [29] |
Shomura A, Izawa T, Ebana K, et al. Deletion in a gene associated with grain size increased yields during rice domestication. Nature Genetics, 2008, 40(8):1023-1028.
doi: 10.1038/ng.169 pmid: 18604208 |
| [30] |
Takano-Kai N, Jiang H, Kubo T, et al. Evolutionary history of GS3, a gene conferring grain length in rice. Genetics, 2009, 182 (4):1323-1334.
doi: 10.1534/genetics.109.103002 pmid: 19506305 |
| [31] | Wu Y H, Zhang B, Cao W H, et al. The ethylene receptor ETR2 delays floral transition and affects starch accumulation in rice. The Plant Cell, 2009, 21(5):1473-1494. |
| [1] | 王若鹏, 吕伟, 刘文萍, 文飞, 韩俊梅, 刘霞霞. 不同栽培模式对芝麻产量及土壤水热状况的影响[J]. 作物杂志, 2024, (4): 247–252 |
| [2] | 苏小雨, 高桐梅, 张鹏钰, 李丰, 吴寅, 王东勇, 田媛, 卫双玲. 基于主成分分析及隶属函数法对芝麻苗期耐热性综合评价[J]. 作物杂志, 2023, (4): 52–59 |
| [3] | 李丰, 高桐梅, 苏小雨, 魏利斌, 王东勇, 田媛, 李同科, 杨自豪, 卫双玲. 施氮量和种植密度对芝麻光合速率、产量和氮肥利用率的影响[J]. 作物杂志, 2022, (2): 215–221 |
| [4] | 肖钦之, 周立, 邓斌, 滕林娟, 李小慧, 秦凌, 王贵, 甘在德, 卢朝军, 崔国贤. 不同单株留蒴果数对湘烟5号种子产量和质量的影响[J]. 作物杂志, 2022, (1): 227–232 |
| [5] | 吕伟, 任果香, 韩俊梅, 文飞, 王若鹏, 刘文萍. 干旱胁迫对芝麻幼苗生理生化指标的影响[J]. 作物杂志, 2021, (5): 172–175 |
| [6] | 吕伟,韩俊梅,任果香,文飞,王若鹏,刘文萍. 山西芝麻种质资源遗传多样性分析[J]. 作物杂志, 2019, (5): 57–63 |
| [7] | 高树广,徐博涵,赵辉,倪云霞,李伟峰,王瑞霞,徐东阳,杨光宇,刘红彦. 芝麻茎点枯病菌Macrophominaphaseolina纤维素降解酶活性分析[J]. 作物杂志, 2019, (4): 196–202 |
| [8] | 高桐梅,李丰,吴寅,魏利斌,王东勇,田媛,费高亮,卫双玲. 不同灌溉方式对芝麻冠层结构及群体质量的影响[J]. 作物杂志, 2019, (3): 162–167 |
| [9] | 吕伟,文飞,韩俊梅,王若鹏,任果香,刘文萍,乐美旺,孙建. 昆虫授粉对芝麻产量构成因素的影响[J]. 作物杂志, 2018, (6): 124–129 |
| [10] | 赵莉,汪强,林勇翔,张祎. 江淮黄褐土壤芝麻肥效研究[J]. 作物杂志, 2017, (6): 154–159 |
| [11] | 桑利民,徐婧,赵晓清,郭元章,蹇家利,郝小玉,郭久林,高永海,韩建国,徐桂真. 地膜覆盖对不同芝麻品种农艺性状及产量构成因素的影响[J]. 作物杂志, 2017, (6): 140–146 |
| [12] | 吕伟,刘文萍,任果香,文飞,韩俊梅,王若鹏. 不同浓度生根粉对芝麻生长及产量的影响[J]. 作物杂志, 2017, (5): 100–105 |
| [13] | 孙建, 魏星, 乐美旺, 等. 芝麻繁育特性研究Ⅱ:异花传粉媒介的确定[J]. 作物杂志, 2015, (3): 36–40 |
| [14] | 颜小文, 乐美旺, 饶月亮, 等. 高产优质芝麻新品种赣芝10号的选育及其栽培技术[J]. 作物杂志, 2014, (3): 151–152 |
| [15] | 杨三维, 张明义. 晋芝8号新品种选育及其栽培技术[J]. 作物杂志, 2013, (4): 154–155 |
|
||