
作物杂志,2020, 第2期: 20–27 doi: 10.16035/j.issn.1001-7283.2020.02.004
高粱硬脂酰-ACP脱氢酶基因(SbSAD)家族鉴定及不同发育阶段表达分析
赵训超,徐晶宇( ),盖胜男,魏玉磊,许晓萱,丁冬,刘梦,张今杰,邵文静
),盖胜男,魏玉磊,许晓萱,丁冬,刘梦,张今杰,邵文静
- 黑龙江八一农垦大学农学院/黑龙江省现代农业栽培技术与作物种质改良重点实验室,163319,黑龙江大庆
Identification of Stearyl -ACP Desaturase Gene (SbSAD) Family and Their Expression Analysis at Different Developmental Stages in Sorghum
Zhao Xunchao,Xu Jingyu(),Gai Shengnan,Wei Yulei,Xu Xiaoxuan,Ding Dong,Liu Meng,Zhang Jinjie,Shao Wenjing
- College of Agronomy, Heilongjiang Bayi Agricultural University/Key Laboratory of Modern Agricultural Cultivation and Crop Germplasm Improvement of Heilongjiang Province, Daqing 163319, Heilongjiang, China
摘要:
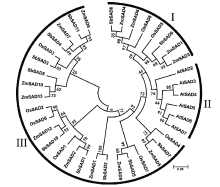
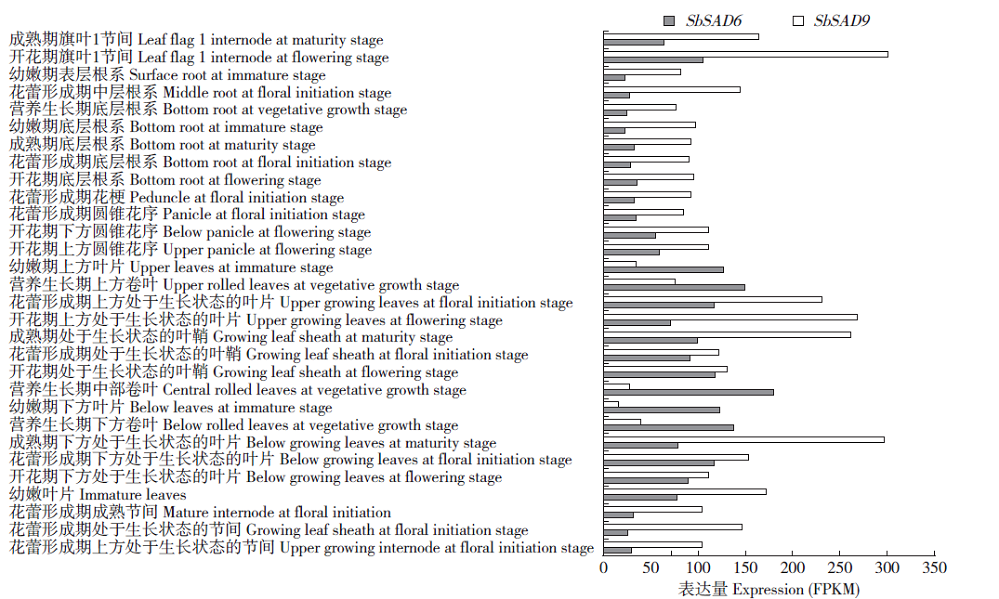
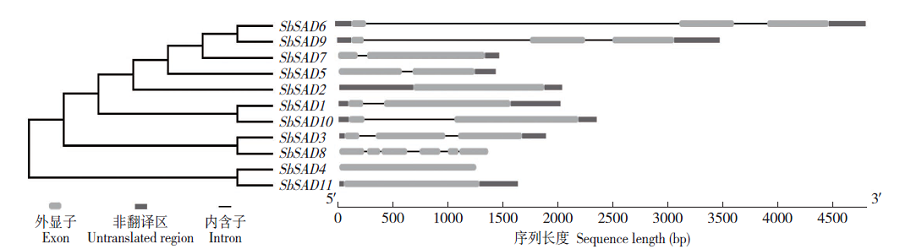
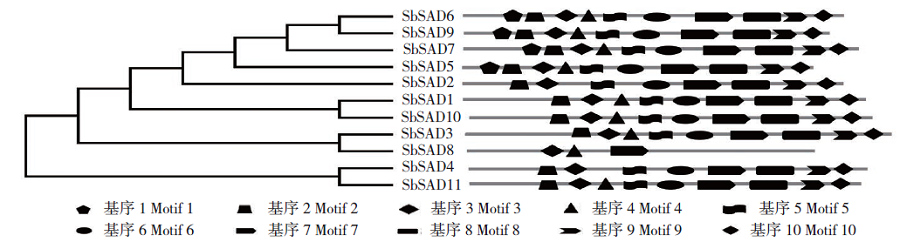
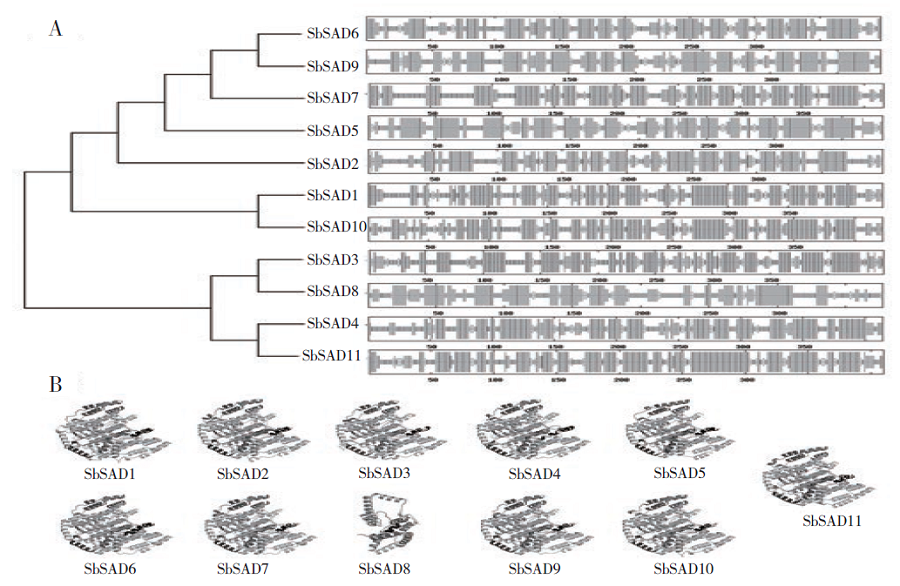
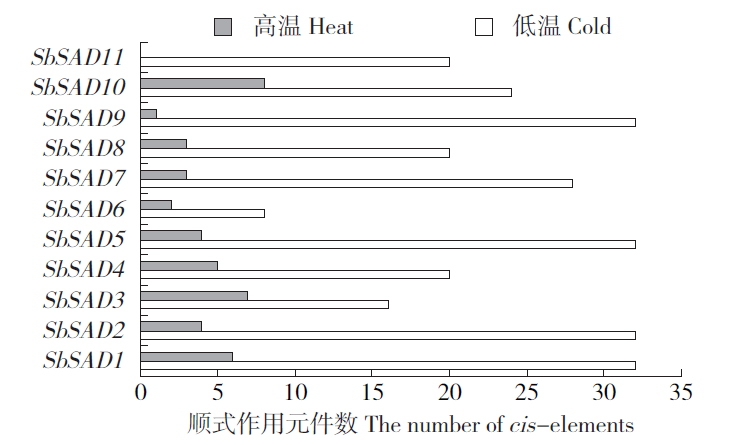
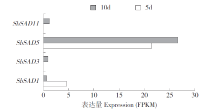
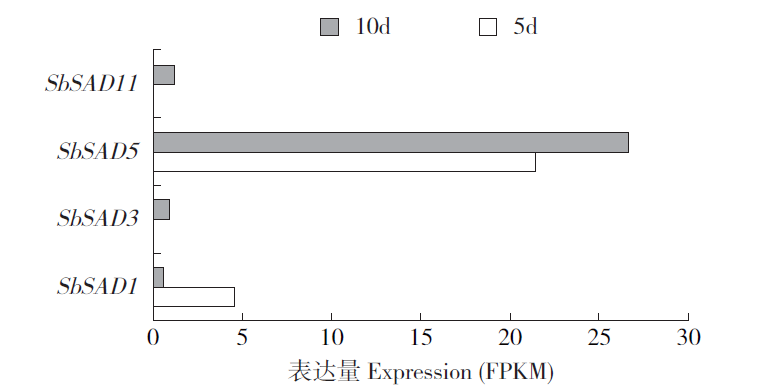
硬脂酰-ACP脱氢酶是形成不饱和脂肪酸的关键酶。利用拟南芥蛋白序列作为比对序列在NCBI和Phytozome数据库进行Blastp同源比对,共鉴定11个SbSADs基因。对SbSAD基因家族进行蛋白特性、进化关系、不同发育阶段表达、基因结构、保守基序、染色体定位、基因二级和三级结构、启动子区的顺式元件及种子发育阶段表达分析,结果显示,SbSADs基因编码区为1 134~1 290bp,其编码蛋白的氨基酸数为377~449,分子量最大为48.0kDa,最小为39.8kDa,等电点为5.26~8.76;根据系统发育树,将SbSAD基因家族分为3个亚族,其中亚族I在不同组织发育阶段表达量较高;11个SbSADs基因不均等地定位在高粱1、2、3、4、6、7和10号染色体上;SbSADs蛋白二级结构均以α-螺旋和无规则卷曲为主,三级结构预测显示,除SbSAD8外,其余SbSADs的蛋白三级结构高度相似;11个SbSADs基因的启动子区与低温胁迫相关的元件数量高度富集。SbSAD5基因在种子发育5和10d均有较高的表达量。







| [1] | 杨士春 . 甜高粱籽粒中脂肪酸含量的气相色谱分析. 中国粮油学报, 2012,27(11):100-104. |
| [2] | 张国琴, 葛玉彬, 张正英 , 等. 高粱抗旱研究综述. 甘肃农业科技, 2018,56(6):67-72. |
| [3] | O'Byrne D J, Knauft D A, Shireman R B . Low fat-monounsaturated rich diets containing high-oleic peanuts improve serum lipoprotein profiles. Lipids, 1997,32(7):687-695. |
| [4] | Carrillo C, Del M C M, Roelofs H , et al. Activation of human neutrophils by oleic acid involves the production of reactive oxygen species and a rise in cytosolic calcium concentration:a comparison with N-6 polyunsaturated fatty acids. Cellular Physiology and Biochemistry, 2011,28(2):329-338. |
| [5] | 袁蕊, 敖宗华, 丁海龙 , 等. 高粱中脂肪酸和低分子有机酸气相色谱测定. 酿酒, 2011,38(4):42-43. |
| [6] | Ohlrogge J, Browse J . Lipid biosynthesis. The Plant Cell, 1995,7(7):957-970. |
| [7] | Xuan W Y, Zhang Y, Liu Z Q , et al. Molecular cloning and expression analysis of a novel BCCP subunit gene from Aleurites moluccana. Genetics and Molecular Research, 2015,14(3):9922-9931. |
| [8] | Gonzalez-Thuillier I, Venegas-Caleron M, Sanchez R , et al. Sunflower (Helianthus annuus) fatty acid synthase complex:beta-hydroxyacyl-[acyl carrier protein] dehydratase genes. Planta, 2016,243(2):397-410. |
| [9] | Rodriguez M F, Sanchez-Garcia A, Salas J J , et al. Characterization of soluble acyl-ACP desaturases from Camelina sativa,Macadamia tetraphylla and Dolichandra unguiscati. Journal of Plant Physiology, 2015,178(15):35-42. |
| [10] | Li-Beisson Y, Shorrosh B, Beisson F , et al. Acyl-lipid metabolism. The Arabidopsis Book, 2013,11:e0161. |
| [11] | Kachroo A, Shanklin J, Whittle E , et al. The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis. Plant Molecular Biology, 2007,63(2):257-271. |
| [12] | Ruddle P N, Whetten R, Cardinal A , et al. Effect of a novel mutation in a △9-stearoyl-ACP-desaturase on soybean seed oil composition. Theoretical and Applied Genetics, 2013,126(1):241-249. |
| [13] | Jung S, Tate P L, Horn R , et al. The phylogenetic relationship of possible progenitors of the cultivated peanut. Journal of Heredity, 2003,94(4):334-340. |
| [14] | Knutzon D S, Thompson G A, Radke S E , et al. Modification of Brassica seed oil by antisense expression of a stearoyl-acyl carrier protein desaturase gene. Proceedings of the National Academy of Sciences of the United States of America, 1992,89(7):2624-2628. |
| [15] | Slocombe S P, Cummins I, Jarvis R P , et al. Nucleotide sequence and temporal regulation of a seed-specific Brassica napus cDNA encoding a stearoyl-acyl carrier protein (ACP) desaturase. Plant Molecular Biology, 1992,20(1):151-155. |
| [16] | Shanklin J, Somerville C . Stearoyl-acyl-carrier-protein desaturase from higher plants is structurally unrelated to the animal and fungal homologs. Proceedings of the National Academy of Sciences of the United States of America, 1991,88(6):2510-2514. |
| [17] | McKeon T A, Stumpf P K . Purification and characterization of the stearoyl-acyl carrier protein desaturase and the acyl-acyl carrier protein thioesterase from maturing seeds of safflower. The Journal of Biological Chemistry, 1982,257(20):12141-12147. |
| [18] | Zhang Y, Maximova S N, Guiltinan M J . Characterization of a stearoyl-acyl carrier protein desaturase gene family from chocolate tree,Theobroma cacao L. Frontiers in Plant Science, 2015,6:239-251. |
| [19] | Lightner J, Wu J, Browse J . A mutant of Arabidopsis with increased levels of stearic acid. Plant Physiology, 1994,106(4):1443-1451. |
| [20] | Osorio J, Fernándezmartínez J, Mancha M , et al. Mutant sunflowers with high concentration of saturated fatty acids in the oil. Crop Science, 1995,35(3):739-742. |
| [21] | Craig W, Lenzi P, Scotti N , et al. Transplastomic tobacco plants expressing a fatty acid desaturase gene exhibit altered fatty acid profiles and improved cold tolerance. Transgenic Research, 2008,17(5):769-782. |
| [22] | Byfield G E, Xue H, Upchurch R G . Two genes from soybean encoding soluble Δ9 stearoyl-ACP desaturases. Crop Science, 2006,46(2):840-846. |
| [23] | Murata N, Ishizaki-Nishizawa O, Higashi S , et al. Genetically engineered alteration in the chilling sensitivity of plants. Nature, 1992,356:710-713. |
| [24] | Kodama H, Horiguchi G, Nishiuchi T , et al. Fatty acid desaturation during chilling acclimation is one of the factors involved in conferring low-temperature tolerance to young tobacco leaves. Plant Physiology, 1995,107(4):1177-1185. |
| [25] | Orlova I V, Serebriiskaya T S, Popov V , et al. Transformation of tobacco with a gene for the thermophilic acyl-lipid desaturase enhances the chilling tolerance of plants. Plant and Cell Physiology, 2003,44(4):447-450. |
| [26] | Dong C G, Cao N, Zhang Z G , et al. Characterization of the fatty acid desaturase genes in cucumber:structure,phylogeny,and expression patterns. PLoS ONE, 2016,11(3):e0149917. |
| [27] | Tocher D R, Leaver M J, Hodgson P A . Recent advances in the biochemistry and molecular biology of fatty acyl desaturases. Progress in Lipid Research, 1998,37(2/3):73-117. |
| [28] | Han Y, Xu G, Du H , et al. Natural variations in stearoyl-acp desaturase genes affect the conversion of stearic to oleic acid in maize kernel. Theoretical and Applied Genetics, 2017,130(1):151-161. |
| [29] | Shang X, Cheng C, Ding J , et al. Identification of candidate genes from the SAD gene family in cotton for determination of cottonseed oil composition. Molecular Genetics and Genomics, 2017,292(1):173-186. |
| [30] | Merlo A O, Cowen N, Delate T , et al. Ribozymes targeted to stearoyl-ACP △9 desaturase mRNA produce heritable increases of stearic acid in transgenic maize leaves. Plant Cell, 1998,10:1603-1621. |
| [1] | 高杰,李青风,李晓荣,封广才,彭秋. 贵州不同年代糯高粱品种(系)干物质生产及光能利用特性差异分析[J]. 作物杂志, 2020, (1): 41–46 |
| [2] | 范昕琦,王海燕,聂萌恩,赵兴奎,张一中,杨慧勇,张晓娟,梁笃,段永红,柳青山. EMS诱变对高粱出苗及农艺性状的影响[J]. 作物杂志, 2020, (1): 47–54 |
| [3] | 宋健,晓宁,王海岗,陈凌,王君杰,刘思辰,乔治军. SiASRs家族基因的鉴定及表达分析[J]. 作物杂志, 2019, (6): 33–42 |
| [4] | 唐桃霞,王致和,施志国,常瑛,张英英,李彦荣. 不同基因型甜高粱对重金属的吸收规律研究[J]. 作物杂志, 2019, (6): 50–56 |
| [5] | 梁晓红,张瑞栋,黄敏佳,刘静,曹雄. 覆膜与施氮互作对高粱产量及水氮利用效率的影响[J]. 作物杂志, 2019, (5): 135–142 |
| [6] | 王劲松,董二伟,焦晓燕,武爱莲,白文斌,王立革,郭珺,韩雄,柳青山. 不同种植模式对高粱晋糯3号产量和养分吸收的影响[J]. 作物杂志, 2019, (5): 166–172 |
| [7] | 岳琳祺,施卫萍,郭佳晖,郭平毅,郭杰. 谷子角质合成基因对干旱胁迫的响应[J]. 作物杂志, 2019, (4): 183–190 |
| [8] | 高杰,李青风,李晓荣,封广才,彭秋. 贵州省不同年代糯高粱品种(系)农艺性状演变分析[J]. 作物杂志, 2019, (4): 17–23 |
| [9] | 李春宏,陆相龙,张培通,苏衍菁,王仪明,郭文琦,殷剑美,韩晓勇,王立,火恩杰. 防除甜高粱田杂草的除草剂筛选[J]. 作物杂志, 2018, (6): 158–161 |
| [10] | 吕亮杰,陈希勇,张业伦,刘茜,王莉梅,马乐,李辉. 小麦GASA基因家族生物信息学分析[J]. 作物杂志, 2018, (6): 58–67 |
| [11] | 张一中,周福平,张晓娟,邵强,杨彬,柳青山. 高粱种质材料光合特性和水分利用效率鉴定及聚类分析[J]. 作物杂志, 2018, (5): 45–53 |
| [12] | 张瑞栋,曹雄,岳忠孝,梁晓红,刘静,黄敏佳. 氮肥和密度对高粱产量及氮肥利用率的影响[J]. 作物杂志, 2018, (5): 110–115 |
| [13] | 张建华,郭瑞峰,曹昌林,范娜,江佰阳,李光,史丽娟,彭之东,白文斌. 几种茎叶除草剂防除高粱田杂草药效和安全性研究[J]. 作物杂志, 2018, (5): 162–166 |
| [14] | 高杰,李青风,彭秋,焦晓燕,王劲松. 不同养分配比对糯高粱物质生产及氮磷钾利用效率的影响[J]. 作物杂志, 2018, (4): 138–142 |
| [15] | 罗海斌, 蒋胜理, 黄诚梅, 曹辉庆, 邓智年, 吴凯朝, 徐林, 陆珍, 魏源文. 甘蔗ScHAK10基因克隆及表达分析[J]. 作物杂志, 2018, (4): 53–61 |
|